年度別活動報告
年度別活動報告書:2006年度
脊索動物と節足動物の共通祖先を理解する 2-1 オオヒメグモの尾葉形成と体軸形成の分子メカニズム
小田広樹(主任研究員) 秋山-小田康子(奨励研究員)
野田彰子(研究補助員) 春田知洋(大阪大学大学院生)
金山真紀(大阪大学大学院生)
はじめに
系統的に遠く離れていると考えられている脊椎動物とショウジョウバエにおいて、相同な遺伝子が個体発生の類似した現象で働いている例が多数見つかっている1)。代表的な例として、前後軸に沿ったパターン形成を制御する遺伝子や背腹軸形成を制御する遺伝子が挙げられる。しかし、このような類似が脊椎動物とハエに見られる理由が、遠い共通祖先の形質を残しているからなのか、異なる起源から偶然に進化したからなのか、脊椎動物と昆虫が実は系統的に近いからなのか、を客観的に判断できるような状況にはない。動物の初期進化の研究が難しい理由は、主に2つの要因からなる。ひとつは、系統関係を推定することの難しさである。最近になって、脊椎動物に最も近い無脊椎動物であると信じられてきたナメクジウオの系統的位置の変更が提案されたことなどからも分かるように2)、3)、最新の分子系統解析法であっても、高次の分類群間の系統関係を解くことは簡単ではない。第二の要因は、動物門という枠組みを超えて形態や発生を比較することの難しさである。例えば、2つの異なる動物種の形を比較するとき、同じ動物門の動物であれば、体の各部位が互いにどう対応するかを判定するのにあまり大きな困難を感じないだろうが、例えば、節足動物門(例えばハエ)と棘皮動物門(例えばヒトデ)の動物であった場合、お互いの体の部位を対応づけることは難しい。「門」という壁を如何に克服するかは進化発生学のひとつの大きな課題となっている。
門は形態を指標に動物を分類したときに見えてくる枠組みであるが、ゲノムに書き込まれた発生プログラムに基づいて動物を分類することができれば、もう少し別の枠組みが見えてくるかもしれない。現在においてはまだ断片的な知識しかないが、異なる動物門に類似した発生プログラムが見つかる一方、同じ動物門に多様な発生プログラムが存在していることがわかりつつある。それぞれの動物門における保存性と多様性をゲノムの情報に基づいて把握することが、動物の統合的な分類体系を構築することにつながるはずである。そこで私たちは、このような研究の方向性を意図し、ショウジョウバエと同じ節足動物門に属しながら、ショウジョウバエから系統的に遠く離れているオオヒメグモを用いて、ゲノム情報に基づいた比較発生学を展開している。ショウジョウバエにおいて蓄積された知識とオオヒメグモの実験動物としての利点を生かして、節足動物門における保存性や多様性に関する知見を効率良く得ることができると考えている。
本研究は将来的に動物門を超えて比較することを目標としており、それを可能にするような初期胚発生の現象に重点を置いている。本年度はオオヒメグモの尾部領域の発生を中心に解析を進めた。形態的な側面だけをみても、ハエ胚とクモ胚の後端部の発生様式には大きな違いがあり、この違いを分子レベルで理解することが節足動物の胚発生の多様性を理解する上で重要であると考えた。
結果と考察
(1)デルタ-ノッチシグナルは尾葉形成に必要である
昨年度までの解析により、ショウジョウバエのデルタの相同遺伝子がツイストの中胚葉での発現に10時間ぐらい先行して将来の尾部領域(原口の周り)の表面上皮細胞層で散在的な発現を示し、parental RNAiによってデルタをノックダウンすることによって、尾葉形成に異常が生じることを明らかにした。本年度はさらに詳細な解析を加え、論文にまとめた(現在投稿中4)。ノッチとサプレッサーオブヘアレスの相同遺伝子のノックダウンでもデルタのノックダウンとよく似た表現型が得られた。分子マーカーの発現解析から、デルタ-ノッチシグナルは原口の周りから生じる中胚葉細胞の数を制限し、尾葉の外胚葉を誘導するのに必要であることが示唆された。
(2)尾葉に特異的に発現する遺伝子の探索
脊椎動物の尾部と節足動物の尾部を比較することを視野に入れて、オオヒメグモの尾葉形成を支配する分子メカニズムの解明を目指した研究をスタートした。正常胚と、尾葉形成に異常が生じるデルタRNAi胚の間の遺伝子発現の差を検出することによって、尾葉に特異的に発現する候補遺伝子を見出すことができるのではないかと考えた。そのために、 EST解析によってこれまでに蓄積してきた配列情報から40merのオリゴDNA配列(プローブ)を12,000種類デザインし、in situ合成方式のマイクロアレイを作製した。そして、尾葉形成期の正常胚と同じステージのデルタRNAi胚からtotal RNAを精製した。異なる蛍光色素で標識した相補的RNA を作製して、競合ハイブリによるマイクロアレイで比較解析を行った。遺伝子発現の差を期待通りに検出できているかどうかを評価するために、コントロールとして9つの遺伝子から21個のプローブ配列をそれぞれ3スポットずつマイクロアレイにあらかじめ組み込んでおいた。コントロール遺伝子のデータから、マイクロアレイで確かにコーダルやオルソデンティクルの発現差を検出できていることがわかった(図1)。そこで、 シグナル強度の比[デルタRNAi胚]/[正常胚]が0.6以下のプローブ配列をポジティブと判定し、42個のクローンを候補として選び、さらにそれらすべてについてin situハイブリダイゼーションを行うことにより、尾葉または中胚葉で特異的に発現するクローンを3つ特定した(図2)。今後はRNAiによりそれらの遺伝子の機能を調べていきたい。
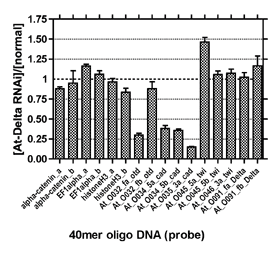 |
図1. マイクロアレイ解析:各種コントロールプローブにおける[At-Delta RNAi胚]と[正常胚]のシグナル強度の比
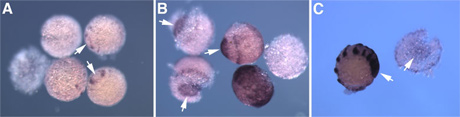 |
図2. in situハイブリダイゼーションによって尾葉または中胚葉で特異的に発現することが判明した遺伝子クローン. 矢印は尾部領域を指している.
マイクロアレイ解析に加えて、cDNAサブトラクションによる解析も試みている。尾葉形成期の正常胚からSMART法によりcDNAライブラリーを作製し、それをもとに同じステージのデルタRNAi胚のcDNAを差し引いた。作製したサブトラクションcDNAライブラリーから、1,405個のクローンの5’末端の配列を決定した。これらの配列をクラスタリングしたところ、1,349個のクラスターに分けられた。つまり、重複した配列は非常に少なかった。差し引きの効果というよりも均一化の効果が強く表れたと考えられた。実際に、その1,405個のクローンの中には、発現量が極めて少ないと考えられるコーダルやデカペンタプレジック、フォークヘッドなども含まれていた。当初の意図とは多少異なるが、このライブラリーの解析を進めることで尾葉形成期に発現する遺伝子を網羅的に調べ上げることができると考えている。
(3)オオヒメグモのヘッジホッグとパッチトの初期パターン形成における役割の解析
ショウジョウバエのヘッジホッグとパッチトはセグメントポラリティー遺伝子として知られている5)。ヘッジホッグは分泌タンパク質をコードし、パッチトはその受容体をコードしている。私たちは、オオヒメグモにおいてヘッジホッグとパッチトの相同遺伝子をクローニングし、それらの遺伝子の発現パターンを調べた。ヘッジホッグは最初胚盤のふちで発現し、その後、形成されつつある尾部領域でも発現が始まる。パッチトは胚盤の中心(原口の周り)と胚盤の周縁部で発現している。胚帯期の胚では、ヘッジホッグとパッチトのいずれもショウジョウバエと相同な発現パターンを示した。
次に、ヘッジホッグとパッチトの初期胚における機能を調べるためにpRNAiによるノックダウン実験を行った。ヘッジホッグのノックダウンでは、クムルス(体軸形成に重要であることが知られている細胞集団)は正常に胚盤のふちへ移動するものの、その後の胚外領域の誘導や胚盤から胚帯への転換が起こらなかった。一方、パッチトのノックダウンでは、胚盤形成やクムルスの形成は正常に行われているようだが、クムルスの移動が起こらず、本来背側に生じるはずの胚外領域が胚盤の中心領域に誘導された(図3)。これらの結果は、オオヒメグモのヘッジホッグとパッチトが体節形成よりも前に体軸形成に必要とされていることを示唆している。この発見は、ショウジョウバエとオオヒメグモの体軸形成機構の違いを理解する上で重要な鍵を握っているのではないかと私たちは考えており、現在さらに詳細な解析を進めている。
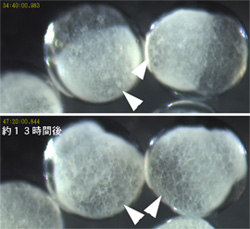 |
図3. パッチトRNAi胚のライブ観察. 胚外領域が胚盤の中心付近にできてしまう(矢印).
(4)シグナルシーケンス・トラップ法によるオオヒメグモ分泌及び膜タンパク質の探索
オオヒメグモの初期胚発生では、これまで私たちが明らかにしてきたように、ソグやデカペンタプレジック、デルタ、ノッチなど細胞の外で働く分泌因子や膜分子が重要な働きをしている6)、7)。一方、ショウジョウバエでは、転写因子の分布が大きな貢献をしている。この違いは、クモなどの鋏角類の胚発生には調節的な側面が多く見られ、ショウジョウバエの胚発生はモザイク的であることと関係していると思われる。私たちはオオヒメグモ胚で働く分泌因子や膜分子をコードする遺伝子を網羅的に探索するために、酵母を用いたシグナルシーケンス・トラップ法によるスクリーニングを開始した。この方法は、シグナルシーケンス領域を欠いたインベルターゼ突然変異株を、アンチマイシンA存在下、ブドウ糖非存在下(ショ糖存在下)でも生育できるようにレスキューする配列、すなわちインベルターゼのシグナルシーケンスの代替となる配列を探索するものである。オオヒメグモ後期胚由来のcDNAライブラリーのスクリーニングにより、確かに、膜/分泌タンパク質をコードする遺伝子が単離されている。現在、さらにスクリーニングを進めている。
おわりに
これまで私たちの研究はショウジョウバエの知識に依存した形で進めてきたが、本年度の研究で新たな道が開けたと言える。RNAi解析、EST解析、マイクロアレイ解析などを組み合わせることで、他の動物種の知識に頼らずに、オオヒメグモの発生現象に関わる遺伝子を効率よく探索することができるようになった。この方法論を生かして、節足動物の初期胚発生を支配する新しい分子システムを明らかにすることができれば、ショウジョウバエや他の動物との比較を通して、節足動物の祖先的な発生を理解するための手がかりをうることができるであろう。