



観る
僕の研究室から出る論文の決定的な証拠写真の多くは今でも僕が撮っています。図版は学生さんが作って、先生が論文全体をまとめるという研究室が普通で、逆は珍しい。構造は普遍だし、幾何学や写真での表現が好きなので自信もある。NatureやScienceなど評価の高い学術雑誌の表紙になったことも何度かあります。表紙は、研究内容と個性的な表現の両方から評価されるものですから、僕としてはこれがとても嬉しいし、皆の心にも残るだろうと思っています。
分野の違う人も含めて、写真を見た誰もが、一目で僕らが発見したことをわかってくれる、そういう像を撮りたいと思い、超薄切片から立体を心にイメージしながら、何時間も、何日も探すのです。一度、見つかってしまえば誰でも探せます。科学者にとって、現象を見つける方法はいろいろありますが、僕の場合は、多様な生物材料の中から誰も使っていない研究目的に最適な材料を探し出し、顕微鏡でまず細胞を観るという方法をずっととってきました。既存のモデル生物では顕現してない興味ある現象は多々ありますからね。



雑誌の表紙を飾った写真。

電子顕微鏡とともに。
遊ぶことの大切さと顕微鏡との出会い
1941年、東京の牛込生まれ。戦時中は長野の小諸に疎開しましたが、戦後すぐに戻り、それからほとんど都心で暮らしています。最初の記憶は、公務員を辞めた親父がはじめた甘味処であんみつを食べている風景です。親父は世話好きで、困っている人や地域の発展のためにせっせと無償で働いていました。その後もいろいろな商売をやったけれど、いつも同じ。店は歳末には福引所、秋祭りには神酒所に変身です。父のそんな姿を見てサービス精神を学んだんじゃないかな。福引きで一等が出た時に、坂の上から下まで鐘をならしながら、「一等~、一等~」と言って走るのが僕の役目でしたし、家族全員祭り好き。祭りの日には神社の花火が朝の10時くらいにシュルシュルと上がり、空でドーンとなる。胸がドキドキ、ドキドキしてね。小学校は午後臨時休校でした。神輿が10台位連なると勇壮で、大人神輿を担いだり、1日に5回くらい神社に行って夜店を覗いてました。
工作が好きで器用だったので図工の時間は生き生きしていましたが、それ以外の勉強や運動は全てダメ。先生に怒られても懲りず、何回廊下でバケツを持って立たされたことか。ガキ大将で、学校が終わると近所の駄菓子屋に集まった仲間を何人も引き連れ、隣の小学校の連中と小さな“公園”を挟んでよくケンカをしました。実はその時のケンカ相手と再会したんですよ。僕が東大の理学部付属(日光・小石川)植物園の園長をしていた時、親睦会で挨拶してくれた理学部長の益田隆司教授が、実はこの近くの“公園”でと話し始めたんです。小さい頃から敵は力より弁がたつなと思っていたんだけど、やっぱりその通り。いつの間にか上にいた訳(笑)。昆虫採集もやったけれど、外堀での鮒やエビ釣りの方が好きだったな。釣り好きは一生続いて、今は友釣りに凝ってます。
4年生の頃、ラジオ作りに夢中になった。配線を間違えてショートさせ家中を停電にしたりしながら完成させたけど音がでない。原因を考えているうちに不眠症になっちゃった。凝り性なんです。自分でもこれはまずい、そこで体を動かした方が良いなと思って柔道を習い始めました。
実はこの頃顕微鏡にはじめて出会ったんです。お祭りの夜店で買った安い顕微鏡で、信州味噌に生えた白いカビを見ていたら、動きまわる虫が見えた。ミクロの世界で予想外なものに出会った驚きというのは今でも忘れられません。多分ダニだったんだろうと思います。その後も中学半ばまで、外堀や池の水をとってきて、その顕微鏡でミジンコをはじめ様々な生物を何時間も見ていました。僕の研究者としての原点はこの時代にある。一方で焼跡の瓦礫を集めた土盛りの中から面白そうな“もの”を見つけるのが好きで得意だったんだけど、マクロとミクロの差はあれ顕微鏡下の世界も同じことです。
逃げた蝶や玉虫を追って神社に忍び込む時には、塀や木登りの得意な奴、追いけるのが早い奴、捕虫網を見事に使いこなす奴、そしてほとんどの虫の名を知っている奴など虫採りにもそれぞれ得意技を生かした分担があった。僕は体が大きく木登りや走るのは苦手だったが、目ざとかったから風で揺らぐ木の葉に隠れた虫を探すのが得意だった。採った昆虫を展翅するより採り方を工夫する方が好きでしたね。こうした子供の頃のちょっとした遊びの延長として、技を生かして新しいものを見つけ、新技術を開発しながら、時には協力しあい謎を解いてゆく今の姿勢があるような気がします。

ガキ大将の頃(前列左)。

中3、仙台への修学旅行にて。

41年誕生。

46年、神楽坂自宅前。道中央は年の離れた姉。ファッション関係の仕事をしていたため、おしゃれだった。この7年後に結核でなくなる。左下本人と妹。

小学校の頃。自宅裏で妹と。

小学校3年生。大好きなお祭りで。最後列、ちょうちんの左。
柔道家から彫刻家、そして生物学へ
都立高校では柔道部を作って柔道に明け暮れました。しかし講堂館には練習しても練習しても勝てない相手がいました。投げられまいと用心して腰を低く構えても、いつの間にか投げられている。凄い得意業をもつ奴がいるもんですよ。研究でも得意技は必要だと思いますね。そんな生活の中で高校2年生の夏休み、ハゼ釣りに行って、海に入った途端夏なのに全身が痺れるような悪寒がした。その前に、姉が結核で亡くなっていたこともあってすぐに病院に行きました。結果は陽性、結核で最低でも1年間の入院と言われ、目の前が真っ暗になりましたね。僕も死ぬと思いました。しかし入院生活で徐々に回復しますとどこが痛いというわけではないから遊びに行きたくて、行きたくて。病院の薬って本当に効いているのかなと思って、検査に出す痰に薬をまぜたら結核菌が出なかった。薬は効くと納得しました。実験魂はこんなところにもあったんだなと思いますね。まもなく退院できて、高校生活に戻りましたが、もう柔道はできない。
仲良しの友人に絵描きの息子がいて、美術の予備校に通っていたので、僕も絵や彫刻をやりたいと思って夜間部に入りました。楽しかったけど、数ヶ月振りに会った彼の瑞々しく輝くようなダビテ像のデッサンに衝撃を受けたのを今でも鮮明に覚えています。まったくレベルが違う、好きだけではダメ、この時も努力にも限界があることを強烈に思い知らされました。それ以後一度も絵は描いていません。柔道もダメ、絵や彫刻もダメ、仕方なく家業を手伝いながらぶらぶらしていました。さすがに親が心配して、外で体を動かす仕事が良いだろうと、測量のアルバイトを探してきてくれました。体を動かすと気持ちも前向きになるんですね。そこで、好きな生きものを顕微鏡でとらえられる生物学に進もうかと思って、近所の悪童仲間と一緒に予備校へ通い始めました。短期間でしたが一生分の受験勉強をしましたね、この時は。
生物の基本は増えることなので、細胞分裂のしくみを知りしたいと思っていました。ところが、そういう勉強ができる大学は少ないことがわかったのです。医学部にもいくつか受かったけれど、結局、ウニを使った細胞分裂の研究で世界的に活躍されていた団勝麿先生に教わりたいと思って、都立大学を選びました。1962年です。

大学時代。小田原に鮎釣りへ。

釣り部の皆と渓流釣りへ

釣ったヤマメと。

かかった!

59年、病院の屋上で柔道部の仲間と。

仲間が帰ると絶望感に襲われた。
電子顕微鏡と出会う
団先生の発生学の授業は3年生になると受けられるので、昼だけでなく夜間部にも出席して軽快な講義を聞いていました。先生は独創的な研究をしなさい、そのためには研究に最適な材料を選び、新しい独自な方法を開発しなさいと教えて下さった。実習の時間には、カエル一匹渡されて、文献を読まずにどんな小さいことでもいいから発見しなさいとか。勉強嫌いの僕の研究姿勢はこれで決まりました。独自に考え最初に発見すれば勉強をしなくてすむ。
この頃、三崎の臨海実験所での実習でウニの卵割をスケッチしたノートに、分裂中の2つの割球の間に矢印をつけて、これは何だ?と書いてある。ウニの発生を観察したら、ほとんどの学生は形ができ上がっていくダイナミックな細胞分化に興味を持つものだけれど、僕は細胞が増える基本的なしくみに興味をもっていたので、分裂の時のくびれに注目したのです。その後、研究の対象は細胞から“細胞内の細胞”であるミトコンドリアや葉緑体へと移りますが、ずっと「くびれ」にこだわって見続けてきたのが僕の研究生活だったな。
二重らせんモデルに続き、ATGCの並びが暗号になってタンパク質が作られることがわかり、生物学に革命が起きていることは学生でも実感しましたね。生命現象を分子、とくに遺伝子で解き明かさなくてはならない。細胞分裂を制御するのも遺伝子に違いないけれど、遺伝子は分裂中は染色体の中にある。これはどういう意味なんだろうと不思議でしたね。そこで、染色体内の遺伝子の正体を知るのが先決と思って、卒研では電子顕微鏡を使って染色体内の遺伝子を観察しました。大学院からは東大の田中信徳先生の細胞遺伝学研究室に移り、染色体数がn=3と少なくて観察し易いキクの一種クレピスを使って染色体の複製機構を調べました。ミクロオートラジオグラフィーミクロオートラジオグラフィー細胞内の放射線で標識された物質を写真乳剤に感光させ光学顕微鏡で同定する方法。という方法で調べたら、3本の染色体は小さい方から大きい方へ、1本の染色体内では末端から動原体の方向に向かって複製が進むことがわかりました。 次に未知だった間期核間期1細胞周期は分裂期と間期に分かれる。間期はG1期、S期(DNA合成期)、G2期からなる。の染色体挙動を観察したいと思いましたが、方法がなかったので超高分解能の電子顕微鏡オートラジオグラフィー法電子顕微鏡オートラジオグラフィー法細胞内の放射線で標識された物質を写真乳剤に感光させ電子顕微鏡で同定する方法。を開発しました。必要なら自分で技術を開発するんだと教えこまれていますからね。この方法により間期の染色体DNAのダイナミックな弛緩と凝縮を明らかにしました。この時撮った写真にはヌクレオソーム構造ヌクレオソーム構造DNAは核内に裸の2重らせん構造としてではなく、タンパク質とともに規則的に折り畳まれて入っている。真核生物のDNAに共通してみられる、DNAの折り畳み基本単位構造。がハッキリ写っている。この構造をもっと追求していればヌクレオソームの発見者になったのに。もったいないことした。見えてないんですよ、写っているけど。同じようなことは幾つかありました。遺伝子の塩基配列を電子顕微鏡下で決定しようとグアニンに特異的に結合する金属を使ってみたり、核小体の機能を調べようと今でいうin situハイブリダイゼーション法を考案したりと、先駆的な実験を行っていましたが、もう一歩ネバリが足りなかったんだな。大分経ってから外国人によって開発され、今世界中の研究室で使われてますね。
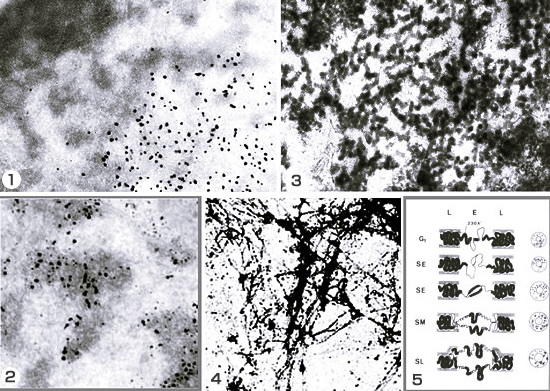
1: 開発した超高分解能電子顕微鏡オートラジオグラフィーで、間期核のDNA合成領域を見る。DNA合成部位を示す銀粒子が分散染色質(明るい)に現れている。
2: 上のから10分間経過したもの。銀粒子は、分散染色質から凝縮染色質へ移行している。
3: 分散染色質の構造を見ると、細いDNA分子が太い繊維となって、高次に組織化されているのが分かる。それらはさらに集まって、凝縮染色質になる。
4: 分散染色質の部分をよく見ると、繊維は球状の小さな顆粒の連鎖であることがわかる。後にこれがヌクレオソームと呼ばれるようになる。
5: 71年博士論文でまとめた、DNAが染色体の中で早期に複製される部分(E,中央)と晩期に複製される部分(L)の細胞周期における構造変化モデル。23nmの繊維をソレノイドと考えれば、現在のモデルに近い。
博士課程の終わりには、いわゆる70年大学紛争、そして大学改革と学生運動が盛んになり、僕は代表者として教授との論争の先頭にたっていた。そんな生意気な学生だったけど、博士論文発表会の後、いつもは対立している古谷雅樹先生が君の研究はユニークで面白いと声をかけて下さった。教育者ってこういうものなんだと思いましたね。今も心に残っています。
70歳を越えた両親が東京に留まってくれと頼むので、都立アイソトープ総合研究所に就職しました。配属先が放射線安全課で、所長から基礎研究を止められた。この時は、課長が説得してくれたおかげで倉庫を片付け、ヒト染色体への放射線の影響についての研究を始められました。ここで原子炉と関係が深いトリチウム水が染色体に異常を起こすことをはじめて見つけ、世界的な話題となりました。今では放射線の管理規定に盛り込まれています。
もうちょっと自由に研究したかったので、岡山大学理学部の武丸恒雄教授の助教授に思いきって応募しました。実は、その直前に岡山での講演会の帰りに、旭川に鮎が大挙して遡上しているのを見てしまったのですよ、また釣り熱がでちゃったんだなぁ。もちろん、それが岡山への応募の第一の理由じゃありませんよ。でも鮎も大いに魅力だった。

三崎の臨海実験所で。手前。
都立大学の臨海実習にて
1964年三崎臨海実習でのスケッチ。ウニの細胞分裂像。娘割球間のブリッジのところを、これは・・・?と示している。以後、細胞とオルガネラでこの部分の構造を追求し続ける。右はウニの受精後、翌日の原腸胚中期とプリズム幼生期の個体。

都立大電子顕微鏡室にて。奥様と酒井文三先生と。

大学院時代。手前から2人目。一番手前は東江昭夫氏(東大教授)。左端は磯野克己氏(神戸大名誉教授)。

72年。岡山の旭川で。鮎の遡上に感動。
ミトコンドリア核の発見と蛍光顕微鏡との出会い
73年に岡山大学に移り、染色体研究を再開しました。細胞周期の特定時期に染色体が弛緩し、それがDNAの転写や複製を誘導したので、この誘導物質を単離したいと思いました。しかし、植物やヒトの細胞では細胞分裂は同調して起こらない。太田次郎先生(お茶の水女子大学)が真正粘菌の変形体は多核なので核分裂を完全に同調化させることができると言ったことを思い出し、粘菌を頂きました。しかし研究室には振とう培養器がないので時々手で振りながら培養したりしていた。学会発表に間に合わせなければならなくなった。簡単な実験を思いつきました。スライドガラス上に粘菌を置き、匙で潰し、シャーレにはった水の上に落とせば核が壊れ、DNAが遊離して活動中の遺伝子が見えるだろうと思ったのです。まだ岡山大学には電子顕微鏡がなかったので、作った試料を東大まで運び、観察しました。大量のゴミの中に、小さな核のような構造を見つけた。これがミトコンドリアの核(DNAとタンパク質の複合体)だったのです。ミトコンドリアや葉緑体のDNAは細菌と同じように、むき出しで入っているといわれていたので、ミトコンドリアDNAが電子密度の高い棒状構造をとっているのに驚きましたよ。粘菌が食べた細菌じゃないかと言われましたけれど、自分ではミトコンドリアの新しい世界を拓けると確信しましたね。そこで細胞核染色体の研究をすっぱりとやめ、ミトコンドリアを“最小の細胞モデル”としてとらえ、これで核の構造と機能、分裂と遺伝の研究をする決心をしました。これが今日まで続くオルガネラ研究の原点です。
岡山大学にも電子顕微鏡が入ったので、粘菌のミトコンドリア核の研究はどんどん進み、ミトコンドリアが、核分裂しながら分裂・増殖していることを発見しました。
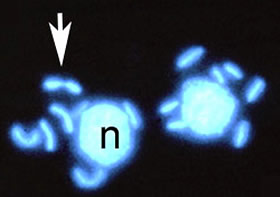
2つの粘菌アメーバ細胞の細胞核とミトコンドリア核を示す蛍光顕微鏡写真。 DAPIで染色すると細胞核(n)とミトコンドリア核は青白色の蛍光を発する。矢印は分裂中のミトコンドリア核を示す。
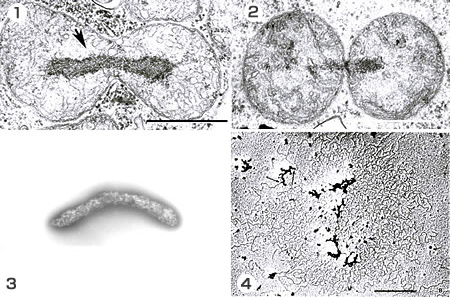
1: 粘菌では、ミトコンドリア核が電子密度の高い部位として電子顕微鏡で観察されるという優れた特性があった。そのため核分裂を伴う(矢印)ミトコンドリアの分裂が観察された。
2: 核分裂後、細胞質分裂に相当するミトコンドリオキネシスが起きる。2つの娘ミトコンドリアの中間のブリッジのところに小さなリングが。しかしはっきりしない。
3: 単離したミトコンドリアの核。
4: 左の単離したものをトリプシンで処理すると、大量のDNAが遊離してくる。
分裂といえば、核分裂だけでなく細胞質分裂があるわけで、ミトコンドリアにもそれ(ミトコンドリオキネシス)があるに違いないと考え、分裂面を観察したけれど、はっきりとしたことはわからなかった。そこで他の生物のミトコンドリアDNAや同じオルガネラである葉緑体DNAも粘菌のように核構造をとり、分裂しているに違いないと考え、その普遍性の証明に力を注ぐ事にしました。
この頃、京都大学の竹内郁夫先生の部屋に遊びに行き、蛍光顕微鏡下で細胞性粘菌の将来胞子を作る細胞が、かなり初期から分化していることを示す鮮明な像を見て、見たいものに蛍光物質をつければ、それだけを選択的に見られるという新しい技術に感動しました。以後それにのめり込むわけです。
岡山は自然が豊かで輝いていましたし、伝統文化の薫りの高い家や社も多く、土地の人は親切で、はじめて東京を離れた遊び人には夢のようなところでした。北は鳥取、南は牛窓周辺の島々までと、しょっちゅう釣りに出掛けました。でかける時間がない時、大学の敷地内を流れる川で釣っていたら、藤茂宏先生に叱られた。先生が父親にそっくりで、、、。でものんびりとした良い時代でした。
岡山生活4年目の77年、神谷宣郎先生(大阪大学)から電話で呼びだされました。先生とは以前、部屋のセミナーに招いて頂いた時にお会いしたきりだったのですが、停年後に岡崎の基礎生物学研究所に移ったので、一緒に研究をしないかとのお誘いでした。岡崎の近くにも良い釣り場がありますよなどと言われ、逆に僕が釣られてしまいました(笑)。

74年、武丸恒雄教授の御家族と。
(前列左から本人、藤茂宏教授、
コルチン教授、武丸教授)

75年、最初の学生達
(左から日詰雅博愛媛大教授、本人、河野重行東大教授)

77年、瀬戸内海で釣り大会。
前列中央本人、左隣は環境ホルモンで活躍している井口泰泉氏。(基礎生物学研究所教授)

77年、学生と富士登山。
「細胞三核説」の提唱とオルガネラの遺伝のしくみ
基礎生物学研究所はまだ建設途中で所帯も小さく、新しいことをやるのだという意気盛んで、活気がありました。本格的にやろうと思い、ミトコンドリア核の普遍性を証明するために、原生生物原生生物アメーバのような単細胞生物や群体及び子実体をつくる単細胞性の真核生物。の分類の大家、オリーブ教授(ノースカロライナ大学)のところへ留学しました。先生は山と積まれたシャーレを実体顕微鏡で毎日々覗き、馬糞塊に発生する原生生物を探していました。僕は先生が持ってきてくれる原生生物を片端から観察し、ミトコンドリア核の存在は不動となりました。しかし、多細胞生物のミトコンドリアや葉緑体のDNAへと展開するには、どうしても微量のDNAの観察が必要だったので、遺伝子1つまで見える超高分解能蛍光顕微鏡(後にオリンパス光学からK型として市販)を開発し、研究所の横にある古池から毎日水や藻を採ってきて、何百もの生物の細胞を観察しました。子供の頃にかえったようで楽しかったですね。その結果、細胞には、細胞核、ミトコンドリア核、葉緑体核の3種があることがはっきりし「細胞三核説」を82年に提唱しました。この説によって、従来見えないものとされていた、ミトコンドリアや葉緑体の超微量のDNAが、確実に見えるものになり、細胞内のオルガネラに対する意識の転換をもたらしました。
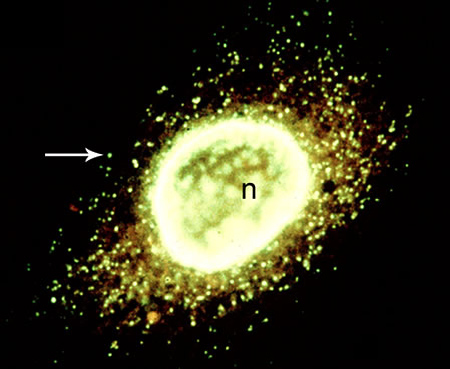
ヒトの細胞核(n)とミトコンドリア核(矢印)を染めたもの。一つの細胞に約2000個のミトコンドリアがある。
オルガネラ核の存在ははっきりしたので、ミトコンドリアと葉緑体の分裂のしくみの 研究を、核分裂と全体の分裂(オルガネラキネシス)に分けて進めることにました。また、超微量のDNAの観察法は出芽酵母の染色体の発見や細胞質(母性)遺伝のしくみの解明にも大きく貢献しました。
ミトコンドリアと葉緑体のDNAは母系遺伝することが遺伝学的にわかっていましたが、そのしくみは明らかではなかった。ところが、オスとメスが同型の緑藻クラミドモナスで、受精30分~40分後、メス由来の葉緑体核はそのままなのに、オス由来の葉緑体核が消える現象を発見しました。これが、オルガネラの母性遺伝のしくみだとすぐに解りました。受精(接合)前後における一連の生化学反応で、オス由来の葉緑体DNAが選択的に分解されることがわかりました。しかし、米国の分子生物学者が時間を追って調べてみると、オス由来のDNAが完全に分解するのは24時間後。30分後では葉緑体の核は分散しただけで、実際にはDNAは分解されていないのではないかと疑問をぶつけられました。僕は1分子まで見える顕微鏡法で観察しているのだから確かだと言ったのですが受け入れられない。
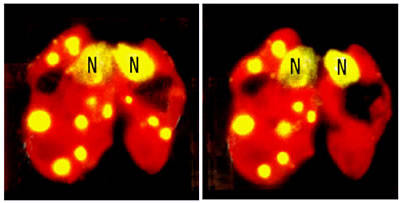
緑藻クラミドモナスの母性遺伝をとらえた接合子の蛍光顕微鏡像。左から右へと30分~40分で右側の雄性配偶子の葉緑体の核が見えなくなる。Nは細胞核。
この頃、研究所も大きく変りました。初代の桑原万寿太郎所長が停年、2代目の金谷晴夫所長は病死、そして84年3代目所長として岡田節人教授が就任しました。僕も長く激しい教授人事を終えて、神谷先生の後任教授になったばかりで緊張気味。教授会で、岡田所長が黒岩さんにたてつくと柔道でぶっ飛ばされるなどと、独特の岡田節にユーモアを交えて話してくれて、緊張から解放されました。
細胞学者が現象を1つの細胞内で見ているのに対し、生化学者や分子生物学者は、複数の細胞から抽出した物質の平均、確率を見ています。生化学実験では、何億ものオス・メスのクラミドモナスを混合しますから、接合してDNAが消えているものもあれば、まだ消えていないものもある。接合しない配偶子だってあるわけです。結局この議論は、1個の細胞の分子生物学をやらなければ、いつまでたっても平行線。そこで、レーザー光をピンセットのように使う光ピンセット法を改良し、1個のミトコンドリアや葉緑体を顕微鏡下で自由自在に移動させ、そのDNAを分析できるようにしました。
クラミドモナス葉緑体DNAに大腸菌の遺伝子aadAを導入し、その葉緑体を持つ配偶子をメス、持たないのをオスとして交配させた場合と、この逆の場合をつくります。葉緑体DNAを生体染色し、オス由来の葉緑体核が消えた接合子1個を光ピンセットで分離し、超高感度PCR超高感度PCR一つの細胞から抽出したゲノムDNA
全体から増やしたい領域だけを増やす
方法。でaadA遺伝子の存在を調べた結果、オスの葉緑体にaadA遺伝子を持つ場合は増幅が起こらなかった。つまり、葉緑体核が見えなくなるということはDNAの完全分解を意味していたのです。ここまでくるのに20年かかりましたけれど、これで決着がつきました。観ることの勝利ですね。現在では、母性遺伝のしくみがよくわかってきました。栄養細胞から配偶子になる段階で、メスの細胞質にDNA分解酵素(ヌクレアーゼC)が合成される。この時、メスの葉緑体DNAには保護機構がつくられる。受精後、メス由来の細胞核の少なくとも3つの遺伝子産物が、ヌクレアーゼCのオス由来の葉緑体内への浸入を助ける。こうしてオス由来の葉緑体DNAは分解され、メス由来の葉緑体DNAは残って子に伝達されるわけです。500種ほどの生物を調べた結果、このしくみは植物全般に普遍的に存在し、ミトコンドリアでもほとんど同じしくみが働いていることが分ってきました。

母性遺伝のしくみを分子レベルでみた1個の細胞のPCR像。交配後30分では変化がないが40分後にはメス由来の葉緑体DNAは分解されない(左)が、オス由来葉緑体DNAは完全に分解される(右)。
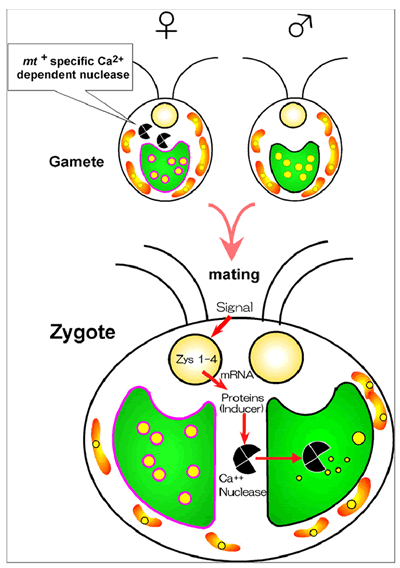
母性遺伝のしくみ
栄養細胞から配偶子がN飢餓によって誘導されると、メスの細胞質内に分子量14万のカルシウム要求性のヌクレアーゼCが合成される。またメスの配偶子内には、このヌクレアーゼに対するDNAの保護機構が発達する。メスとオスの配偶子が接合すると、その接合のシグナルがメス由来の細胞核の遺伝子を活性化する。これらの遺伝子産物はヌクレアーゼCがオス由来の葉緑体に入るのを助ける。こうして入ったヌクレアーゼCは葉緑体のDNAを分解する。

建設中の岡崎基礎生物学研究所の前で。(左端本人。左から4人目が神谷宣郎教授。)
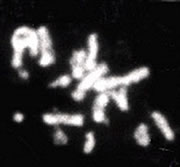
出芽酵母の染色体の発見。16本数えられますか?

ブラウン大学から超高解像度顕微鏡法を習いに来たコールマン教授と。

81年、浜名湖で黒鯛を。カモメと二人きり?

微小管の国際会議にて。
(後列左から5人目本人。前列左から5人目は団勝磨教授、6人目D.メイジャ教授)

81年、遺伝子組換え班会議で富士山を背に。
(左から本人、杉浦昌弘名大教授、榊佳之東大教授、富野士良都立大教授)

粘菌の国際学会にて。粘菌の研究者は大きい?
色素体リングの発見とオルガネラの分裂増殖のしくみ
僕の頭の中には、いつもミトコンドリアと葉緑体の分裂像があります。粘菌ではミトコンドリア核が明瞭でしたので、核を単離して生化学的に調べ、“ミトコンドリアミン”と名付けたタンパク質の解析と核分裂の解析を精力的に進めました。詳細は省きますが、こうしてミトコンドリアの核構造や核分裂のしくみが分子レベルで明らかになり、細胞で言えば細胞質分裂にあたる現象、オルガネラキネシスのしくみを集中して解析できました。
粘菌をはじめほとんどの生物では、1個の細胞に含まれるミトコンドリアや葉緑体の数が数十から数千個と多く、しかもデタラメに分裂するので、分裂像を容易に探せない。そんな時、大先輩の田宮博先生が、53年に光の明暗で緑藻のクロレラ細胞を同調的に分裂させたことを思いだしました。もっと原始的な藻類は、葉緑体、ミトコンドリアがそれぞれ1個で、同調的に分裂させることができるに違いない。そこで新しい研究材料を探しました。草津温泉産の原始紅藻シアニジウムがまさに望みの生物で、細胞核、ミトコンドリア、葉緑体がそれぞれ1個だったのです。細胞分裂の同調化にも何とか成功し、分裂像を追いました。86年、ミトコンドリアのくびれ部分に“構造”を探していた時、葉緑体のくびれ部分の外側にリングが巻きついているのを発見しました。
この頃、東京大学に移り、飯野徹雄、田沢仁両教授の協力でスムーズに研究が進み、88年には葉緑体のリングの普遍性が明らかになりました。リングは色素体分裂リング(PDリング)と名付けました。64年ウニの割球のくびれを見て、これは何だ?と思って以来、くびれの形の本体を追い求めていたので、このリング発見は嬉しかったですね。我ながら執念深いと思います。
このリングは、真核細胞の成立の際に、宿主細胞がオルガネラの祖先である細菌の増殖を制御するために作った装置と考えると、細胞の起源の謎を解くとっかかりになるはずだ。そこからはもうまっしぐらです。
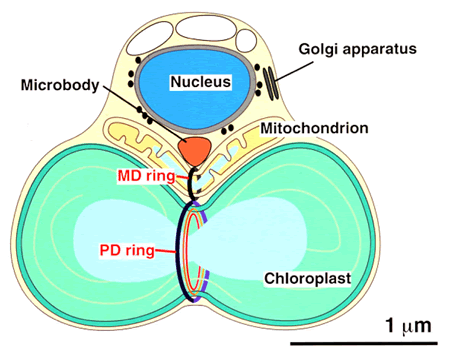
分裂中のシアニジウム類(シアニジュウムとシアニディオシゾン)の模式図。細胞の直径は1-2μ程度で、上から細胞核、ミトコンドリア、葉緑体を1個ずつ含む他、マイクロボディ、ゴルジ体も1個である。
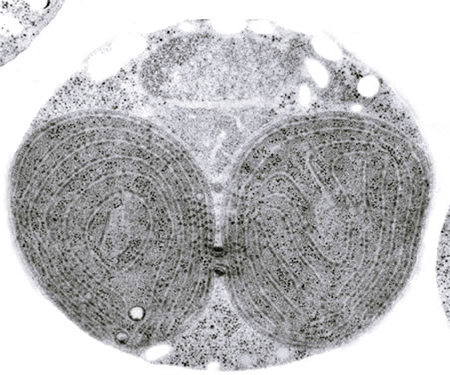
シアニジウムの電子顕微鏡像。上から細胞核、ミトコンドリア、分裂中の葉緑体が観察される。葉緑体の分裂くびれ部分にPDリングの断面が黒くみえる。

87年4月。東大理学部2号館前にて。
20億年前からの贈り物
その後、いくつかの研究室から報告されたさまざまな植物でのPDリングの構造を比較していて面白いことに気が付いたのです。僕が最初に発見したシアニジウムのPDリングが最も大きく、高等植物では最も小さい。進化とともに小さくなり、構造も単純になっているのです。シアニジウムより原始的で単純な生物で、もっと大きなPDリングと粘菌より明瞭なミトコンドリア分裂リングが発見できるかも知れない。世界中を探しました。
92年、原始紅藻シアニディオシゾン(通称シゾン)と出会ったのです。ナポリ大学の温泉藻の分類学者が78年にはじめて報告し、その後ほとんど研究がなされていなかった単細胞の原始紅藻で1回の分裂周期が1生活環。すぐに採集のつもりでイタリアへと向かいました。幸いナポリ大学のピント教授が持っており、少し分けていただいたのですが、その時にはまだ純化されておらず、多くの温泉藻と混合したまま培養されていた。持ち帰って、シゾンを単離し純粋培養しました。真核生物には珍しく、強酸性でかつ45度の環境に生息します。光で細胞分裂を同調化してみると、1個ずつある葉緑体、ミトコンドリア、マイクロボディ、ゴルジ体、細胞核が、順次同調的に分裂し最後に細胞質分裂がおきました。オルガネラの分裂を含む細胞分裂に関する研究のモデル生物として作られたみたいな生物。こんな素晴らしい材料は他にありません。
おかげで、93年にはついにミトコンドリアの分裂面の外側に分裂リング(MDリング)を発見できました。その後の研究で、葉緑体では5重、ミトコンドリアでは4重のリングを使っていることまでわかりました。葉緑体の分裂をまとめると、まず葉緑体の核タンパク質HUによってDNAが核分裂のために組織化され、次に細菌の分裂に使われているのと同じFtsZリングが葉緑体の分裂予定面の一番内側に形成されます。続いて内側、中間そして外側のPDリングが形成され、すぐに分裂面の収縮が起こります。収縮が進み終盤になると、FtsZリングと内側のリングは解体され分裂面から外れます。外側のPDリングは収縮とともに太くなり、分裂のための力を発しているようです。しぼられたPDリングを見ると、丁度、絹糸が巻かれた糸巻きのように見えます。
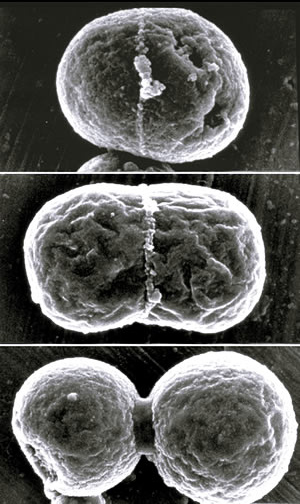
シゾン葉緑体の分裂過程を示す走査型電子顕微鏡像。分裂赤道面の外側に出現したリング(PDリング)は収縮し、太くなり、葉緑体を分断する。

シゾン葉緑体分裂リング拡大写真。分裂の動力は直径4ナノの”夢の糸巻”(PD/MDリング)。
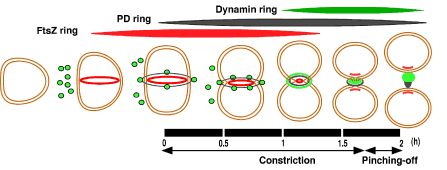
葉緑体の分裂過程を示す模式図
分裂開始前にFtsZリングが分裂赤道面の内側に形成される。続いて外側のPDリングが形成される。収縮が進むと細胞質にあったダイナミンが分裂赤道面に移動し、最初は外側のPDリングの周辺に集合するが、直ぐに外側のPDりングの内側に入り、葉緑体を分断する。FtsZリングは分断直前に分解されるか、娘葉緑体に分配され、消失する。内側と中間のPDリングは示されていない。驚いたことに、これはミトコンドリアの分裂過程と全く同じであった。
直径僅か4nmの細い繊維の糸巻き。こうして葉緑体は分裂・増殖し、光合成によってわれわれの食糧となるでんぷんを生産するのですから、人類の命綱はナノレベルでの力仕事と言える。外側のPDリングが巻きついた葉緑体は古代の壷のようにも見えました。そこで、退官記念の冊子の表紙に「20億年前からの贈りもの」と題して写真を載せたのですが、皆さんわかってくれたでしょうか(笑)。PDリングだけでは葉緑体は分断しません。最後にダイナミンというタンパク質でできたリングが外側のPDリングの内側に形成され、それで2つの娘葉緑体に分断されるのです。この最後のダイナミンのリングは、これまでの観るという手法とは異なる方法で発見しました。僕の得意技ではない、ゲノム解析です。観ることにこだわり、だからこそ独自の発見ができたと自負していますが、これはそれだけでは発見できなかっただろうと思います。生命現象を知るにはできる限りの手法を活用しなくては。

やっぱり釣りが好き。

光ピンセットでオルガネラを移動。超高解像度顕微鏡と。



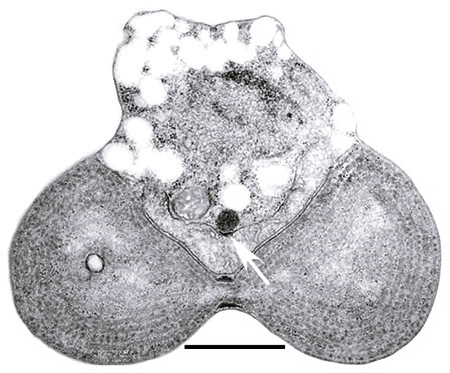
シアニディオシゾン(シゾン)の電子顕微鏡像。上から細胞核、V字型をしたミトコンドリア、そして分裂中の葉緑体が観察される。矢印はミトコンドリアのMDリングを示す。
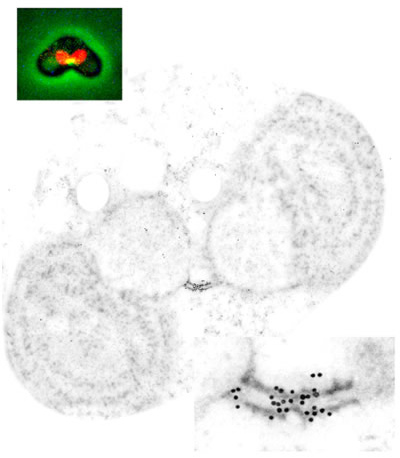
ダイナミンの局在を示す免疫蛍光顕微鏡像と免疫電子顕微鏡像。蛍光像ではミトコンドリアは赤く染められており、その分裂のくびれ部分にダイナミンのシグナル(黄緑)が現れている。免疫電子顕微鏡像ではダイナミンのシグナルを示す金粒子は分裂中の娘ミトコンドリアを繋ぐ“橋”の中央に現れている。右下は拡大図。
シゾンのゲノム解析
PDリングを単離してそのタンパク質から遺伝子を決定したい。しかし解析できるタンパク質は超微量です。微量なタンパク質の解析にはどうしてもゲノム情報が必要だし、シゾンのミトコンドリアと葉緑体のゲノムの全塩基配列を決定していたので、細胞核のゲノム情報が得られれば分裂に関わる全貌が解けると思いました。しかも、シゾンは真核生物の起源に限りなく近いところから分岐した最小ゲノムセットの遺伝子を持つ真核生物である。しかし、停年退官直前で、ヒトに比べたら小さいゲノム(17Mbp)とはいえ、たくさんの人の協力が必要なプロジェクト型の研究を進めるのには抵抗があった。それでもさまざまな協力を得たことやゲノムプロジェクトに必要な得意技を持つ学生達を“発見”できたことも幸いし、ゲノム解析は順調に進み、現在99.9%以上の塩基を読み、完全に繋げた染色体20本に6000弱の遺伝子があることが明らかになりました。シゾンはほとんどの遺伝子が最小セットなので、多細胞生物や酵母では、複数ある遺伝子が一つだけのことが多い。ダイナミン関連の遺伝子は2つあり、それぞれが葉緑体とミトコンドリアの最後の分断に関与していました。ゲノム解析の威力ですね。





東大教授室で。机は団先生が使っていたもの。
尽きない大河ドラマ
ミトコンドリアと葉緑体の分裂・増殖、遺伝の研究は現存の生物がどのように生きているかという問題と、20億年前真核生物がどのように誕生したかという問題とに直接関わっています。ダイナミン遺伝子やPDリング遺伝子は細菌にはありませんから、これらのリングは宿主細胞がオルガネラの祖先である細菌の分裂を制御するために作り出した装置だろうと思います。ほとんどの真核生物のミトコンドリアと葉緑体のゲノムは、自立に必要な遺伝子の90%以上を、細胞核に奪われていますし、母性遺伝によっても支配されています。ミトコンドリアと葉緑体は宿主細胞に寄生した細菌から始まっているとされていますが、宿主細胞核ゲノムは、細菌と共生する際に、細菌から多くの遺伝子を短期間に抜き取り、分裂装置で分裂を制御し、母性遺伝により遺伝子組換えの阻止機構を作動させ、細菌をエネルギー変換器に変えたように思います。
さらに驚くことがわかってきました。ミトコンドリアも葉緑体もほとんど、いや全くと言っていいほど同じ装置、FtsZ、MD/PDリング、ダイナミンリングを使って分裂していることです。MD/PDリングの遺伝子を早く決定したくてしかたがありません。この類似性は何を意味しているのでしょうか。細胞内共生はαプロテオ細菌をミトコンドリアにする時とシアノバクテリアを葉緑体にする時の2度起きたか、あるいはこれらの起源細菌にただ1度だけ起きたのかという問いは大事なものです。いずれにせよ、もっと検証は必要ですが、ミトコンドリアと葉緑体の起源が同じという可能性もあり、そう考えるのも面白いというのが現段階です。
こうした研究データは、未来への重要なメッセージを提示することになるでしょう。今後、二酸化炭素による温暖化、砂漠化、飢餓や人口爆発など、深刻な問題が一気に噴出する可能性があります。こうした問題に共通に関わっているのが植物の保全と生産性の向上です。植物機能の現場は葉緑体なのですから、葉緑体がどのように分裂するかの基本原理の解明は近い将来必ず役立つはずだと思っています。顕微鏡下で小さな小さなミトコンドリアと葉緑体が過去現在未来へと繋がる壮大なドラマを熱演してくれているような気がしてならないのです。
心で観る
科学を芸術にというのが座右の銘ですが、40年かけてそろそろ芸術に近づいてきたかな(笑)。何に限らず真理は単純で美しい。そして新しいものが見えたら次の新しい世界が開けるはずですから、常に新しいものを探していく。研究は本来、創造です。僕は顕微鏡を使って本質の美しさを求め、暴き、描き出してきた。ミトコンドリアや葉緑体はとても小さい。何百、何千というサンプルの中から、伝えたいことの本質を一番あらわしているものを探すのです。これにはセンスが必要で、やはり努力だけでは無理です。顕微鏡は目で観るのではなく、心で観るものです。若い頃、視野にはあったのに、見えなかったという体験を何度かして、それがよくわかってきました。つまり想像していないと出会えない、見えないのです。人は皆それぞれ得意技を持っていますが、僕の得意技は心で観ることでしょうか(笑)。
最近は科学者の顔が見える研究が少なくて残念です。皆、本来得意技がある。現象の発見は心ときめくものですが、真の得意技を持った学生や研究者の“発見”も嬉しいものです。僕も限られた時間のなかで、こうした研究者に智恵をかり、ゲノム情報を顕微鏡と同じように真理を暴く武器として使いながら、オルガネラの分裂と遺伝の全貌を解明したいと思っています。
多くの先輩学生に支えられてきましたが、とりわけパートナーである黒岩晴子博士に感謝しています。これまで彼女が登場しなかったのは、一緒に研究しながら、具体的テーマは少し違うものだったからです。彼女と一緒に卒研をして以来、いつも一緒に研究をしてきました。彼女の興味は一貫して高等植物の重複受精のしくみですが、僕の研究も理解しよい助言をくれます。よき人生の友を得てはじめて一つのことに打ち込めたのだとあらためて思っています。

馬堀海岸にて石川統教授、守隆夫教授の研究室とラボ対抗釣り大会。









