最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。
バックナンバー
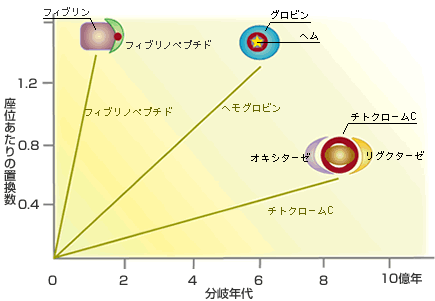
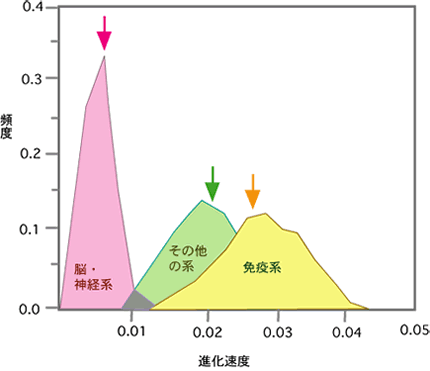
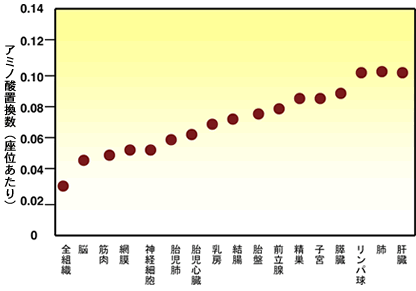
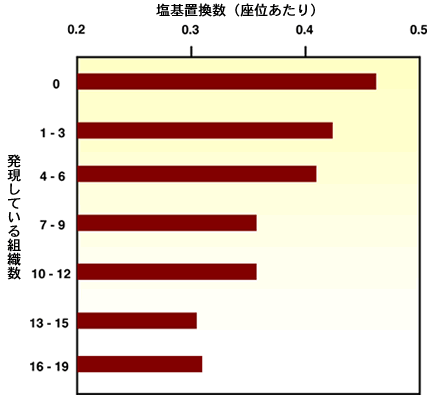
【分子の進化速度は形態と分子の進化を橋渡しできるか】
2005年8月1日
|
閉じる
(日月火水木金土)
10:00-16:30
![]()
最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。
バックナンバー
【分子の進化速度は形態と分子の進化を橋渡しできるか】
2005年8月1日
|