RESEARCH
普遍的な左右対称性のしくみを探る
ハエの消化管から見る
左右非対称な形づくり
動物の外見はほぼ左右対称ですが体の器官には左右非対称性がよく見られます。器官の左右非対称性は遺伝的に決められているので、動物の発生や進化を考える上で重要です。脊椎動物や軟体動物では分子レベルでのしくみは明らかになりつつありますが、細胞、器官レベルではよくわかっていません。節足動物であるショウジョウバエの後腸(消化管)は胚発生の初期には左右対称ですが、発生がすすむにつれて反時計回りにねじれて左右非対称になります。東京理科大学の前田礼男さんは、器官が非対称になるしくみの一端を分子・細胞レベルで解き明かしました。今までに見つかっていない新しい左右非対称性の形成機構であり、更なる研究が期待されます。
1.動物界にみられる左右非対称な形態
動物の形は大雑把に見ると左右対称である。しかし、カタツムリや貝類の渦巻き状の殻、ヒラメやカレイの目の位置、シオマネキの大小のハサミなど非対称なところもたくさんみられる。さらには、外見上は左右対称だが、内部は非対称ということも少なくない。人間でも、心臓、胃、膵臓は左側、肝臓は右側に偏って位置している。左右非対称性がどのようにして起こるかという課題は、古くから多くの生物学者の興味を引いたが、長い間解かれなかった。そんな中、1990年代半ばに、左右非対称性に関する分子機構の一端が明らかになったのである。からだの左側に特異的に発現する遺伝子、 Nodal、 Lefty、 Pitx2が発見され、左右非対称性に関する研究が飛躍的に発展した。しかし、この遺伝子情報に基づいて、左右非対称な形態が形成される機構については、よくわかっていなかった。そこで私達は、モデル生物として長い歴史をもつショウジョウバエを用いて、左右非対称な形態が生まれる機構を明らかにする研究に取り組んだ。
2.左右非対称性を制御する遺伝子を求めて
ショウジョウバエが遺伝学の研究材料として使われ始めたのは今から約100年も前のことで、多くの知見が蓄積されている。そして、ホメオボックス遺伝子(註1)の発見に代表されるように、ショウジョウバエでの研究が、ヒトを含む多くの動物に普遍的な原理の発見につながることが少なくない。
ショウジョウバエでは、胚発生時の消化管で、まず左右非対称性が見られる(図1)。私達は、これを制御する遺伝子を見つけ出すために、左右非対称性に異常を示す突然変異体系統を人為的に作り出し、それに関わる遺伝子を探索した。そのような遺伝子は複数存在するのだが、そのうちの一つであるMyosin 31DF (Myo31DF) 遺伝子のホモ接合体胚では、後腸と中腸の後半部の左右非対称性がほとんどの個体で反転することがわかった(図2)。この遺伝子は、細胞骨格の一つであるアクチン繊維と相互作用するミオシンタンパクの一種をコードしている。実は、胚発生過程で左右非対称性が野生型と反転している個体は、成虫まで生存可能、生殖能力も正常であり、成虫のからだの中も高頻度で逆転していることが明らかになった。
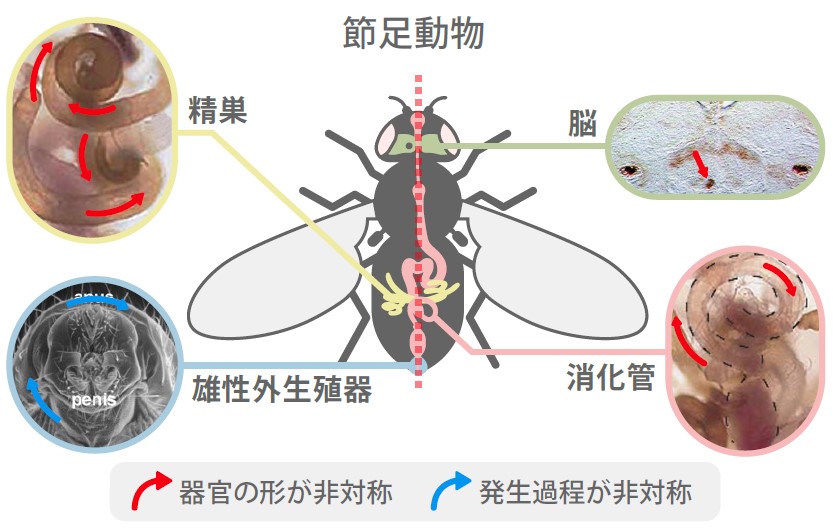
(図1) 節足動物(ショウジョウバエ)の体の中にある左右非対称な器官
ショウジョウバエの脳・精巣・雄性生殖器・消化管などは左右非対称性を示す器官である。なかでも消化管が発生の過程で最初に左右非対称性を示す。
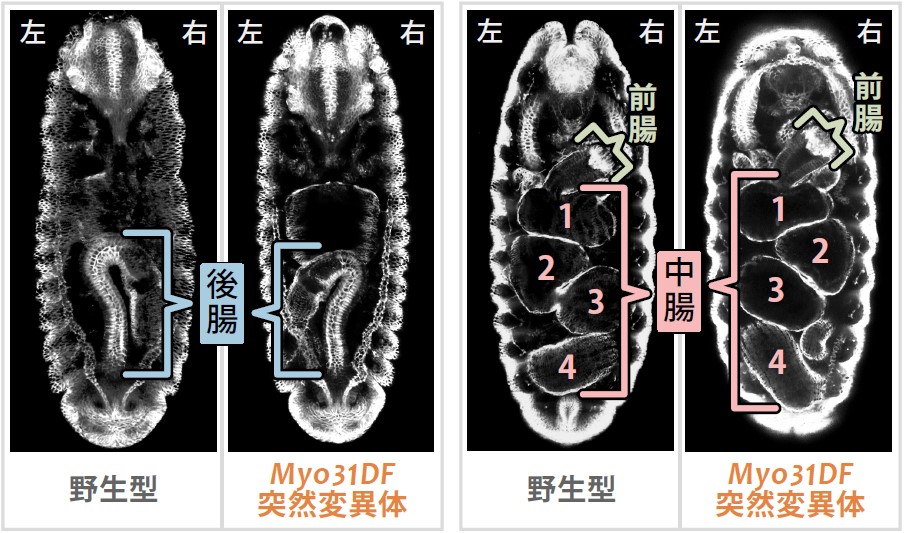
(図2)Myo31DF突然変異体において反転した左右非対称性
Myo31DF突然変異体においては、胚消化管の後腸と中腸の後半部が、ほぼ全ての個体において反転する。
次に問うべきは、 Myo31DFなどの遺伝子が、消化管の形態を左右非対称に変化させる仕組みである。そこには、細胞レベルの何らかの変化があるはずであると考えた。 私達は、細胞の詳細な観察とコンピュータ・シミュレーションを用いた新しいアプローチとで、この謎に取り組んだ。
(註1)ホメオボックス遺伝子
動物の形態形成などを制御する遺伝子のグループで、DNAに結合する180塩基対のホメオドメインを共通にもつ。
3.キラルな細胞形状が左右非対称性な臓器形態を形作る
ショウジョウバエ胚発生時の後腸は、最初、からだの正中線(註2)上に左右対称な状態で位置している。それが発生が進むにつれて、後方からみて反時計周りに90度ねじれて左右非対称になる(図3)。この時、細胞はどのようになっているのだろう。細胞分裂や細胞死はほとんど起こっていないので、私達は、上皮細胞の「形」の変化が後腸の反時計周りのねじれを誘発しているのではないかと推測した。
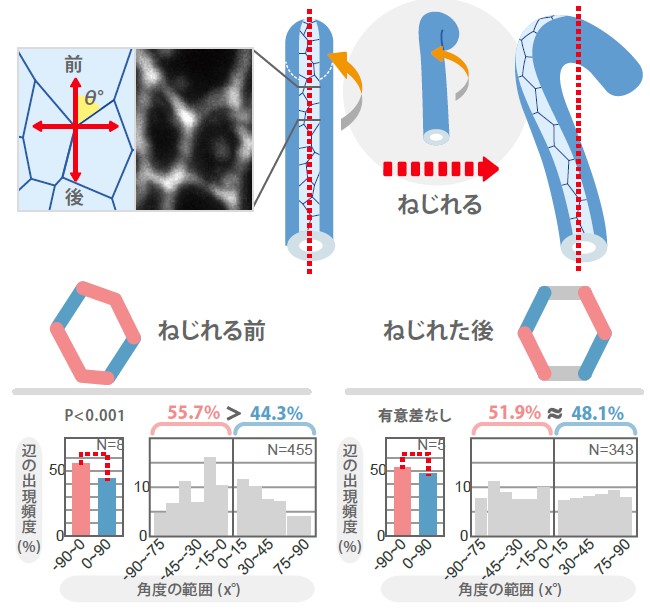
(図3)後腸上皮細胞のキラルな細胞形状
ねじれる前の後腸上皮細胞の形状を調べたところ、その形が左右非対称であることが明らかになった。一方、ねじれた後の細胞形状は、ほぼ左右対称な形である。
そして、上皮細胞の形の測定をつぎのように行った。後腸上皮細胞の頂端面(後腸上皮の管の内側)に注目し、後腸の前後軸を基準(0度)に、それと2つの細胞の接触面(細胞辺という)との間の角度を測定し、その分布を調べた(図3左上)。その結果、右へ傾いている細胞辺(0度から90度の範囲にある細胞辺)と左へ傾いている細胞辺(0度から-90度)の出現頻度に偏りがあることがわかった。これは、後腸がねじれる前に、細胞の「形」は左右非対称になっていることを示唆している。この事実を発見した時、頭に浮かんだのが、「キラル(chiral)」という言葉だった。キラルとは、ある物体が相互に鏡像になっている関係をいう。右手と左手はキラルの代表例である。この実験では、後腸上皮細胞の頂端面の形に注目して左右非対称な「形」を発見したが、実際の後腸上皮細胞は頂低極性をもった立体構造(註3)をしている。これは、その形が左右非対称であり、その鏡像と重ね合わすことのできない性質を持っていることから、私達は、後腸がねじれる前の左右非対称にゆがんだ細胞の性質を「Planar Cell-shape Chirality (PCC)」と名付けた。ねじれ後の後腸上皮細胞では、左右非対称なゆがみは観察されなかった(図3右)。また、後腸の左右非対称性が逆転するMyo31DF突然変異体でPCCを調べたところ、細胞の左右非対称なゆがみも逆転していたので、ねじれる前の後腸上皮細胞の形状が、後腸のねじれる方向を決めている可能性が考えられる。
野生型とMyo31DF突然変異体の後腸でみられる左右非対称な細胞形状が形づくられる過程を明らかにするために、細胞接着分子であるE-Cadherin (E-Cad)をコードしているDE-Cadherin (DE-Cad)遺伝子に注目した。DE-Cad突然変異体では、後腸の左右非対称性がランダムになっており、後腸がねじれる前の細胞形状は、ほぼ左右対称であることがわかった。ここから、E-Cadの細胞辺への分布の偏りがPCCを形成しているのではないかと考え、細胞辺におけるE-Cadの分布を免疫蛍光染色で可視化し相対的な分布を算出したところ、野生型では分布が左右非対称になっていた。一方、Myo31DF突然変異体では、その分布の偏りが逆転していたのである。この結果から、私達は、図4のような仮説を立てた(図4)。野生型では、最初、後腸上皮細胞の初期の左右極性にしたがってE-Cadが左右非対称に分布する。その後、MyoIDの機能によってE-Cadの左右非対称な分布が逆転する。E-Cadが力学的に細胞接着面に作用することで、それぞれの細胞辺に左右非対称な力がかかり、正常なPCCが形成されるのである。一方、Myo31DF突然変異体では、MyoIDが機能しないために、初期の左右極性にしたがったE-Cadの左右非対称な分布が維持されることになる。その結果、野生型とは逆の左右非対称な収縮力がかかり、反転したPCCが形成されるのではないだろうか。
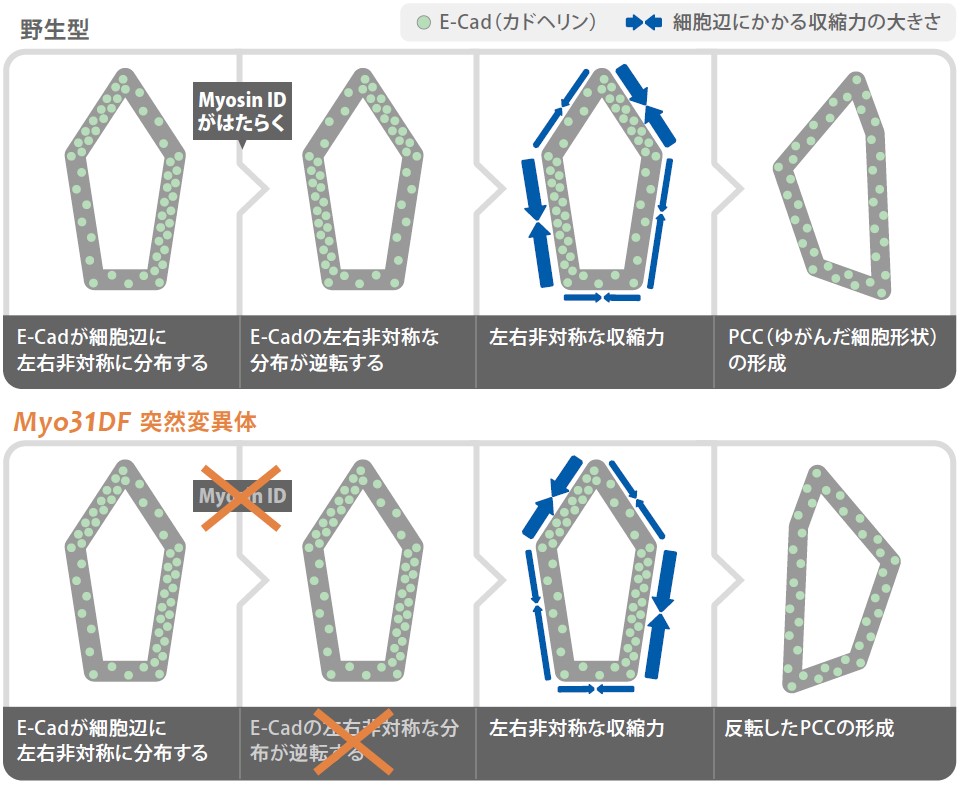
(図4)MyoIDとE-CadによるPCC形成機構の仮説
野生型では、MyoIDの機能によって、E-Cadの左右非対称な分布が逆転するため、結果的に、左右非対称にゆがんだ細胞形状、PCCが形成される。Myo31DF突然変異体では、初期の左右極性にもとづいたE-Cadの分布に依存するので、野生型の場合とは逆のPCCが形成される。
(註2)正中線
生物体の前・背面の中央を頭から縦にまっすぐ通る線のこと。(青色が正中線)
(註3) 頂低極性をもった立体構造
特定の方向に沿ってその両極端に相対応する異なった性質を極性という。後腸細胞の形状は管の内側(頂端側)と外側(基底側)で性質が異なる立体構造をしている。
4.コンピュータ・シミュレーションを用いた後腸のねじれの再現
ここまでの研究から、私達は、各細胞でE-Cadが偏った分布をすることによってPCCが形成されると推測した。さらに、生体内では、ねじれた後の後腸上皮細胞の左右非対称な歪みが観察されないので、その後のPCCの解消が後腸のねじれを引き起こすのではないかと考えた。しかし、このような単純な機構で、多数の細胞から構成される後腸のねじれを説明できるだろうか? そこで、仮説を検証するために、後腸上皮細胞を想定した多角形パターンをコンピュータ上に構築した。
このモデル後腸上皮で、細胞辺にかかる収縮力に左右差を与えたところ、実際の後腸上皮細胞でみられるようなPCC様のパターンがみられた。次に、PCC様のパターンを示すモデル後腸上皮細胞が安定した形をとるように、収縮力の左右非対称性を解消したところ、実際の後腸と同じようなねじれが再現できた。このコンピュータ・シミュレーションによるモデル検証の結果から、E-Cadの細胞辺における分布の偏りによって生じるキラルな細胞形状が、多数の細胞から構成される組織の形態変化を誘発する可能性を示すことができた。
5.動物界にみられる細胞レベルのキラリティ
ショウジョウバエの後腸が左右非対称になるのは、基になる細胞がキラルな性質をもっているからであるという意外な研究成果は、一見すると左右差のないその他の細胞もキラルな性質をもっているのではないかと考えさせる。実は、近年、この可能性を示唆するような研究成果がいくつか報告されている。たとえば、ヒト好中球の培養細胞では、中心体と核を結んだ軸の左側に仮足を伸ばす傾向があることが報告されている。さらに、ゾウリムシなどの多くの原生動物のからだにもキラリティが存在する。これらのことを合わせると、実は、キラリティは細胞の基本的性質の一つであり、多細胞生物に見られる左右非対称な形態は、細胞のキラリティの結果として見られるものなのかもしれない。
私達は、ショウジョウバエの後腸に注目して、左右非対称にゆがんだ細胞形状「PCC」の形成にMyo31DF遺伝子とDE-Cad遺伝子が関与していることを明らかにした。ただし、PCCの形成にはその他多くの未知の遺伝子が関わっていると考えられる。私達は、すでに、ショウジョウバエ消化管の左右非対称性に関与する遺伝子をいくつか見つけ出しているが、それらの左右非対称性形成に関する機能はほとんど未解明である。これらの遺伝子の機能を解析し、PCC形成のより詳細な分子機構を明らかにしていきたいと考えている。今後も、この小さなショウジョウバエをパートナーとして、動物に普遍的なメカニズムの理解という大きな期待を胸に、研究に取り組んでいきたい。
謝辞
本内容の中心となった研究論文の共同筆頭著者である谷口喜一郎博士、シミュレーションに多大な協力を頂いた安藤格士博士、そして本研究の遂行を支援してくださった松野健治教授、本研究に携わった松野研究室のメンバー全員に心から感謝致します。
引用文献
図1 脳写真:Nature 427,605-606(2004)
図1 生殖器写真:Development 130,2397-2406(2003)
前田礼男(まえだ・れお)
2007年東京理科大学大学院基礎工学研究科博士後期課程修了。博士(工学)。現在同大学基礎工学部生物工学科助教。ショウジョウバエを用いて左右非対称性形成のメカニズムを研究している。