

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
分子進化をダーウィンの進化論に当てはめる
2015年12月1日
生命誕生を考えるとき、前回述べた熱水噴出孔に形成される泡のような小胞ほどよくできた偶然はないように思う。うまい具合に大きさも細胞に近く、小孔で他の小胞や外界ともつながっているし、原理的にアミノ酸、核酸、脂肪酸、ペプチド、ヌクレオチドなどを持続的に合成する条件も整っている。さらに、アセテートやメタンの合成過程でエネルギーを発生させることができ、熱勾配やpH勾配のおかげで高分子を特定のコンパートメントに濃縮することもできる。
ただ、どんなに細胞に似ていても、これは生命ではない。チムニー内の小胞では、熱力学平衡に対抗するため、不断にエネルギーを外界から導入して(ここではH2勾配、CO2の還元反応、熱などがこれに相当する)熱力学的平衡から離れた状態を維持できる開放系、すなわちイリヤ・プリゴジンが散逸構造と名付けた非平衡状態が実現できている。しかも、この状態を何万年にもわたり維持することができる。しかし、ここで起きていることの全ては生命誕生に必要な条件ではあっても、生物特有の性質とは無関係だ。非平衡の熱力学系ができただけで、生命誕生までにはまだまだ大きなギャップが存在している。一般的に創発と呼ばれてきた質的な転換が必要で、まずこのギャップで何が起こったのか大きな枠組みを考える必要がある(図1)
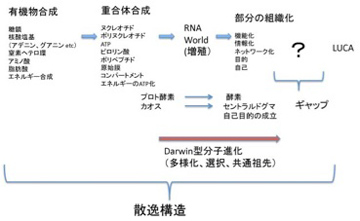
図1 最初の生命(LUCA:Last Universal Common Ancestor)で起こったと思われることを列挙した図。
前にも述べたが、英語では生命をOrganismと表し、生物が「組織化されている」ことを強調する。これはフランス語(Organisme)、イタリア語・ラテン語の(Organismo)も同じだ。ところが日本語やドイツ語では「生物」(Lebenswesen)と表すため、有機化学と生命誕生の間に横たわるギャップの本体が、「組織化される」過程であることを、いまひとつ実感しにくい。18世紀、デカルトの機械論では生命を理解できないと反旗を翻した自然史運動の中心ビュフォンを代表とする先人たちが目指したのが有機体論だったことを思い起こそう。その後、生物が組織化された存在だという特徴は、生物には「自然目的」が内在しているように見えるという特徴へと読み替えられ、19世紀生物学の重要なテーマとしてダーウィン進化論の誕生まで受け継がれていく。
組織化するメカニズムはそのまま訳すとオーガナイザーであり、シュペーマンが使用して以来、発生学で頻繁に用いられる用語だ。分子を生命へと組織化するという、実際には何も理解できていないメカニズムをとりあえず生命のオーガナイザーと呼んでおこう。もちろんこの本体については全く分かっていないことから、ここではオーガナイザーという言葉を、この過程を理解できていないことを表す目印として使う。と言うのも、間違うとオーガナイザーという概念は、神を始めとする超越的力による組織化と結びつけられる危険がある。事実、ダーウィン以前の生物学では、オーガナイザーとは確実に神の持つ超越的力のあらわれと考えられていたし、ダーウィン以後も「エラン・ビタール(生命力)」などの超越的力は、繰り返しオーガナイザーと結びつけられてきた。これはオーガナイザーという言葉自体に、最終目的を知り、そのために部分を組織化する主体という意味が付きまとうからだ。オーガナイザーと表現したときは、常にわかっていないことの印であることを思い起こし、超越的説明を導入しないように気をつけながら進む必要がある。
オーガナイザーを理解するため、過去の生物学の歴史にもう一度目を移してみよう。18世紀生命を組織化するオーガナイザーは、生命の「自然目的」と読み換えられていたが、この自然目的を科学的に初めて説明したのがダーウィンの進化論だった。重要なことは、進化論が超越論的説明を完全に排除してこの自然目的を説明している点だ。従って生命誕生までのオーガナイザーを考える時、ダーウィンの進化論は大いに参考になる。
ではダーウィンの進化論は生物の持つ「自然目的」をどう説明しているだろう?この説では、目的を実現すべく進化したように見える種(例えば高い木の実を食べる為に首が長くなったキリン)も、最初から小さな違いが集まった集団の中の多様性として存在し、この小さな差が他の個体と比べた特定の環境下での生殖(増殖)優位性により自然選択されただけだと説明する。すなわち、進化では目的として提示される結果が最初から存在していたという逆転の発想だ。
次回以降順に取り上げるが、ダーウィンの扱った生命誕生以後の進化と、それ以前の分子進化には様々な違いが存在する。しかし、結果が先に多様性として存在し、それが選択されるという図式は分子進化にも当てはめられるのではと考えている。すなわち、生命誕生と非平衡熱力学系の間に、無数の中間状態(分子の種類とそれらの関係性)が存在し、多様な分子の集まったカオス状態の中から、特定の中間状態が選ばれる過程が繰り返されるうちに、最後に全生物に共通の祖先(LUCA)が誕生すると考えることができる。
この分子進化のダーウィン的理解に必要なのは、1)誕生までに発生した中間段階、2)生命誕生に至るまでに必要な分子や中間段階多様性を発生させる力、そして3)分子から生命への過程に働く自然選択力になる。今回からこの3つの問題を順に考えて行く。
ダーウィンの進化論では共通祖先の概念は、出し手のいない情報を扱うための素晴らしいアイデアになっている(これについても次回以降議論する)。このおかげで、現存の生物を単一の原始細胞まで遡って構想することが可能になる。同じように分子進化でも、単純な構造(例えばディペプチドやディヌクレオチド)から複雑な構造へと分子進化が起こり、最終的にLUCAに集まったのだろうと推察はできる。もっとも単純な有機体をメタンとアセトンとすると、知る必要があるのは、このもっとも単純な有機体構造から合成されてきた、自立生命を支えるために必要なミニマムな分子の構造リストだろう。
進化論では、生物の複雑化を示す中間段階の証拠として化石の存在が指摘されている。しかし残念ながら、化石に相当する中間段階の遺物は、38億年前に起こった生命誕生までの過程を示すほど多くは残っていないだろう。再現できず、記録もない過去の分子を研究するには、現存する分子を基礎にして過去の分子を推論するための方法が必要になる。現在この目的で広く行われているのが、DNAの配列の差に基づいて系統関係を推定し、その共通祖先を推定する分子系統学だ。しかし、DNA情報が成立していない生命誕生前にはこの手法は役立たない。生命誕生までに生まれた中間状態や、分子構造の複雑化について考える分子進化は、有機分子の構造だけを指標として進化を考える必要がある。ゲノム配列を使うことに慣れた私たちには大変に思えるが、例えばダーウィンの進化論は、形態の観察だけから生まれてきたことを考えると、できない話ではない。
幸い構造解析の終わったタンパク質はすでに10万を突破している。アミノ酸配列からタンパク質の構造を自動的に予測することはまだ難しいため、タンパク質をその形態から分類しようとCATHと呼ばれる分類が1993年より行われている(図2)。
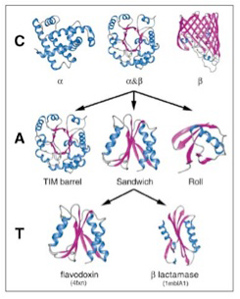
図2 Orengoらの論文で行われたタンパク構造の階層化。Cはclass, Aはarchitecture, TはTopology 。これによりタンパク質の構造の複雑化の過程を、情報ではなく、形質として研究できる。(Orengo et al, Nucl. Acids Res. (1999) 27 (1): 275-279.より転載)
この方法では、タンパク質をClass,(類) Architecture(構築), Topology(接続形態)、そしてhomologous superfamily(相同スーパーファミリー)に階層化している。全てのタンパク質は、αヘリックス類、βシート類、そして両方が合わさったαβ類に分けられる。この3類がそれぞれ多様な変容を遂げることで異なる形態を持つ構造が生まれる。タンパク質の複雑化が進むと、同じ類に属していても、形が大きく違った構築を作ることができる。これがarchitectureだが、図1ではαβ類から派生しうる3種類のarchitectureが示されている。こうしてクラスより複雑な構造の多様なarchitectureを一定数抽出することができる。
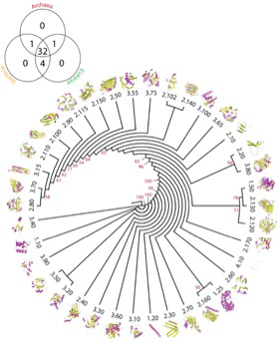
図3 Caetano-Anollesらにより提案されているarchitectureの系統図。ここでは38種類のArchitectureが区別され、構造、配列、機能を総合した推計方法で系統樹が描かれている。左端のベンダイアグラムはそれぞれのArchitectureが古細菌、原核生物、真核生物にどう分布しているかが示されている。このレベルの構造はほぼすべての生物に存在し、従ってよりLUCAの持っていたタンパク質に近い構造だと想像できる。(Bukhari and Caetano-Anolles, Plos Computational Biology, 9:e1003009, 2013より転載)
図3は、Caetano-Anollesらが、構造の決定されたタンパク質からarchitectureを抽出して系統関係を調べた図だが、重要なことはほとんどのarchitectureが古細菌、原核生物、真核生物のすべてに存在することだ。すなわち、このような構造リストは、生命誕生に必要なミニマム分子のリストに近いと考えられる。一方、様々なarchitectureが結合したtopology, homologous superfamilyなどのより高次な構造になると、存在する生物は徐々に限られていく。
このように、タンパク質やRNAの構造の階層性、細胞内での機能、そしてアミノ酸配列総合して進化過程を記述しようとするCaetano-Anollesらの努力は、生命誕生後の進化を新しい観点から見るだけにとどまらず、例えばATP合成や、アミノアシルtRNA合成酵素のような生命の基本に関わる酵素のもっとも単純な構造を推定する方法になる可能性がある。
もちろんこのもっとも単純な機能タンパクリストが完成しているわけではないが、次回からは、LUCAに集まったこの分子群のリストが完成したと仮定して、分子多様性を生み出す力、generator of diversity (GOD)について考えたいと思う。