

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
コミュニケーション・伝達・記録
2016年10月3日
前回まで、ゲノム、フェロモン、クロマチン構造が媒介する情報と進化で生まれた新しい情報について見てきた。これらの情報を、伝達、コミュニケーション、記録という3つの情報機能について見直してみよう。
伝達、コミュニケーション、そして記録は情報が可能にする重要な機能だ。ゲノムはこれらすべての機能を併せ持っている。新しく生まれた情報には伝達機能は存在するが、フェロモンには記録という機能はなく、クロマチン構造を媒介とする情報はコミュニケーション機能を持たない。前回述べたが、フェロモンとクロマチン構造はゲノムの持つ情報を補完するように別々に誕生し、真核生物になると統合された情報システムへと発展する。そしてこの統合から生まれた新しい可能性が、多細胞動物の誕生の基盤になる。今回は、情報の記憶とコミュニケーション機能について整理をした上で、多細胞体制が生まれるまでに、生物の情報システムがどう発展したのか整理しよう。
まず、フェロモンやクロマチン情報が必要とされた背景、情報としてのゲノムの限界から見てみよう。
いうまでもなく、細胞が複製できるのは、ゲノムがDNAを記憶媒体として使って生物のオペレーションを指示する情報として働いているからだ。この記録機能と記録の伝達の原理については、DNAが2重螺旋構造をしていることが発見された時、解明されたと言っていい(図1)。
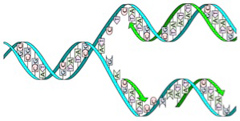
図1 2重螺旋の複製。2重螺旋が一本鎖にほどけ、それが2本の2重螺旋になることがDNAの構造から明らかになり、伝達と記録機能の原理が理解された。(wikipediaより)
ではコミュニケーション機能はどうだろう。複製されたゲノムは2つの娘細胞に伝わる。これは生殖を通しての情報伝搬だが、コミュニケーションではない。
ゲノム同士のコミュニケーションは基本的に相同組み換え機構を介して行われる。私たちの体細胞には、母親からの染色体と、父親からの染色体が存在するが、条件が整うと両方の染色体同士で相同組み換え(図2)が起こり、染色体の部分交換がおこる。これにより異なるゲノム同士のコミュニケーションが可能になる。
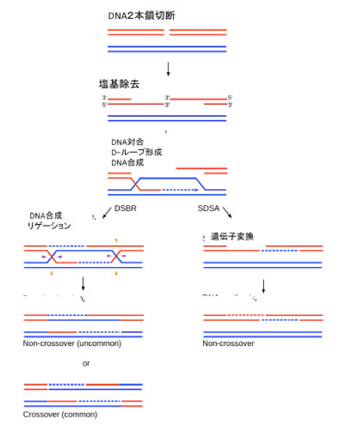
図2:相同組み換えを通した2つのゲノムのコミュニケーション(wikipediaより)
この機構はほぼすべての生物で保存されており、大腸菌から人間まで、異なる個体のゲノム同士のコミュニケーションにはこの機構が使われる。ただ、相同組み換えが起こるためには、DNAの切断が必要で、実際には放射線などによるDNA切断に対する修復機構として進化してきたと考えられる。通常の状態で相同組み換えが起こるためには、DNAを自然に切断する過程が必要で、細胞にとっては危険極まりない。このため多くの生物では、この機構を通したゲノム間のコミュニケーションが高い確率で起こる状況は限られている。例えば、人間では生殖細胞の減数分裂過程に限られ、体細胞で起こる頻度は低い。
このように、ゲノムの記憶、伝達、コミュニケーションの様式を見てみると、全てが生殖サイクルと完全に一体化していることがわかる。単細胞動物では、異なる個体同士が接合するとゲノムレベルのコミュニケーションが起こるのを観察できるが、コミュニケーションの範囲は1対1に限られており、一つのゲノム情報が同時に異なるゲノム情報とコミュニケーションすることはない(図3)
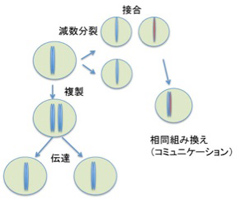
図3: ゲノムには情報が記録されており、複製を通して娘細胞に伝達される。また、接合、ゲノム間の相同組み換えにより、他の個体のゲノムとコミュニケーションできる。ただ、図からわかるようにゲノム情報の伝搬、コミュニケーションは全て一対一で個体(細胞)が対応しており、同時に複数の個体とコミュニケーションしたり伝達することはない。
ではなぜコミュニケーションが必要か、その利点を考えてみよう。ダーウィン進化を現代的に翻訳すると、様々な様式のゲノムレベルの多様化がまず起こり、ゲノムの多様化が形質の多様化として反影されると、この形質の環境適合性を指標に生殖優位性での差異が生じ、環境に適応した個体の比率が高まることで進化が進むと言えるだろう。このゲノムの多様化の一番の駆動力は、DNA複製に必ず伴うエラーだが、もう一段質の高い多様化の駆動力が相同組み換えによる多様化だ。
全くランダムな多様化と比べると、相同組み換えによる多様化は、ゲノムレベルのコミュニケーションを基盤にしている点が大きく異なる。通常これが起こるのは性生殖だが、この場合異なる環境で生きてきた個体がそれぞれ持っているゲノム同士の部分を交換し合う。視点を変えてこの過程を見てみると、その種が分離してから世代を重ねてきた異なる個体のゲノム同士、いわば環境によりすでに「テスト済み」のゲノム同士で情報交換が行われていることを意味する。それぞれの個体のゲノムがその個体が出会ってきた環境を同化していると考えると、この環境との相互作用の歴史が一つのゲノムの中へ再構成されているとみることができる。言葉を変えると、ゲノム間のコミュニケーションにより、より多くの環境との出会いが、ゲノムに同化される。これがゲノム間でコミュニケーションが行われることの重要な役割だ。
この様にゲノムには、情報としての全てが備わっている様に思える(事実それが進化の原動力となってきた)が、特定の生物集団の生存から見た時、ゲノムの対応能力には限界がある。すなわち、ゲノムだけで環境の変化に対応しようとすると、伝達やコミュニケーションが1:1に限られ、空間的にも、時間的にも急激な環境変化には対応できない。この限界を埋めるのが、フェロモンと、クロマチン情報だ。
詳しく検証したわけではないが、フェロモンは、細胞内の分子間相互作用ネットワークが、細胞外へ拡大したものと考えられる。フェロモンは接合の相手を惹きつけ、接合時の方向性(オスメスと考えて貰えばいい)を決める過程にしばしば登場する。これは1:1の個体間相互作用と言えるが、他にもクオラムセンシング(前々回)のように1:1の限界を超え、同時に多くの個体とコミュニケーションをとりあう手段を提供し、全ての個体が環境に対して同じように反応するための情報としても働いている。
この様に、生存環境に関する情報を他の個体と同時的に共有することを可能にしたのがフェロモンの機能だ。フェロモン自体の情報には記録機能がないため、フェロモンのシグナルは一時的な反応を誘導して終わるか、あるいはフェロモンの情報を引き金に、遺伝情報を書き換え(酵母の交配型の変換)や、プラスミドを介したゲノム情報のやりとり(前々回)を行うことで、経験が記録される。このゲノム自体の書き換えの代わりに、クロマチン情報の書き換え機構とフェロモンシグナルをリンクさせることもできる。実際、クロマチン情報の書き換えが、反応の多様性、伝達範囲の大きさ、伝達と記録の迅速性の点で、ゲノムの書き換えをはるかに凌駕しているため、これがその後の生物の主要な伝達様式になっていく。
前回みた様に、クロマチン構造を媒介とする情報は、書き換え、すなわち繰り返して記録が可能だ。このメカニズムが完成するのは、おそらく真核生物以降だが、これに必要な個々のメカニズムは原核生物や古細菌で進化してきた。事実DNAメチル化は原核生物から存在するし、ヒストンと相同性を持つタンパク質も古細菌から認められる。更には、遺伝子の相同性は欠如しているが、DNAと結合してヌクレオソーム構造を形成するためのタンパク質は原核生物にも存在している。
この様なヌクレオソーム構造が広く認められることは、プラスミドの様な小さなゲノムは別として、メガベース以上の大きさのゲノムは裸のままで存在することが不可能であることを示している。実際、バクテリアやアルケアでは、1mmの長さのDNAを1ミクロンの大きさの細胞内に安全に収納している。その上に、収納した全ゲノムをなんども複製することが求められる。ヒストンの様な、DNAをコンパクトに巻き取る仕組みなしには、ゲノムサイズの急速な増大はありえなかっただろう。
真核生物の誕生にはヒストン修飾を基盤とするクロマチン構造制御機構、分裂時の染色体分離に必要なチュブリン、そして形態や細胞分裂を制御できる細胞骨格分子アクチンが必要だが、この3種類のタンパクは同じ分子からできてきたと考える魅力的な仮説がある。すなわち、いずれもリン酸化やアセチル化による修飾を介した細胞の構造変化に関わる。この考えを採用した、真核細胞進化過程について図3にまとめてみた。
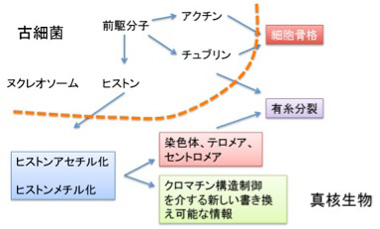
図3:古細菌から真核生物への進化をヒストンから眺めてみる。
ここでは話をヒストンに限るが、最初DNAをコンパクトに折りたたみ保護するヌクレオソーム形成分子として誕生したヒストンは、N末端に集まるリジン残基のアセチル化/脱アセチル化、メチル化/脱メチル化を介する修飾により、単純なヌクレオソームから、各ゲノム領域を構造的に標識するクロマチン構造へと発展した。これにより、テロメアやセントロメアのように、有糸分裂に必要な条件が整った染色体が生まれる。そしてゲノム領域を構造的に標識するヒストンの同じ能力が、遺伝子発現調節に使われるようになったのが、クロマチン情報だ。
春化の例でクロマチン情報は、環境のストレスを、多くの個体が同時に、同じシグナルとして感知し、このシグナルを同じ様に記録することを可能にしている。このようにクロマチン情報は複数の個体が同時に共有できる、ゲノムではなしえなかった情報と言える。
最初環境ストレスに対する反応として誕生したクロマチン情報の書き換え機構に、フェロモンによる細胞間情報共有機構が統合されると(図4)、それ自身では互いに未完成であった情報が補い合って、コミュニケーション、伝達、記録のすべて備わった新しい情報システムが完成する。
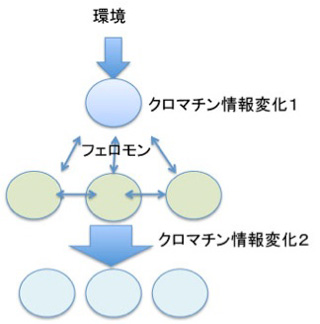
図4:環境ストレスによるクロマチン情報の書き換え機構にフェロモンが組み合わさると、より複雑な細胞間のコミュニケーションが可能になる。
図4で示した構造をよく見てもらうと、ある環境シグナルに対して直接対応する細胞と、その後フェロモンの作用で反応する細胞が別々に描かれているのに気づいてもらえたと思う。これは多細胞体制を想定して描いている。多細胞体制では、個体と環境、個々の細胞と環境、個々の細胞間、そして個体と個々の細胞間の4種類の関係が成立しており、この関係から生じる細胞の変化はすべてゲノムの変化を伴わない、クロマチン情報の変化が背景にある。例えば、個体と個々の細胞の関係には、同時的な情報の共有と、細胞レベルの経験の記録が必須で、これはクロマチン情報とフェロモン情報(細胞間情報伝達機構)だけで行われている。逆に多細胞体制成立から考えると、フェロモン情報とクロマチン構造を介する情報が誕生し、それが統合されることが、多細胞体制誕生の必須の条件であることになる。