

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
記憶と個性
2017年1月5日
ここまで、ゲノム、クロマチン情報、そして神経細胞ネットワークについて、情報と記憶という観点から検討してきた。今回はこれまでのまとめとして、3種類の情報システムを、個性という観点から整理してみようと思っている。
個性というと、「差異、多様性」だけを思い浮かべがちだが、例えばサルと人間の違いを個性とは言わない。個性というとき、あくまでも人間、あるいはサルの個体間で見られる違いについての話だ。したがって、個性という言葉の中には、種としての同一性と個体としての多様性の両方が含まれている。言い換えると、環境に適応するため種としての共通性を維持しようとエントロピーを低下させようとする方向と、ゲノム変異により個体を多様化しようとエントロピーを拡大する方向がぶつかり合うところに個性が成立している(図1)。

図1: ゲノム変異は常に種内のエントロピーを上昇させ、これに対し自然選択はエントロピーを低下させる方向に働く。この結果、種という制限の中で多様性が生まれ、個性が形成される。
おそらくダーウィンほど(ウォレスもだが)、この個性の理解が進化を理解する鍵であることがわかっている人はいなかっただろう。種という制限の中で、次の世代に伝えることが可能な個性が自然に生まれることを確信できないと、自然選択説はでてこない。
人間に個性があるように、あらゆる動物にも個性があることを一旦認めれば、産業革命下の弱肉強食社会を目の当たりにしていたダーウィン時代の都市のインテリにとって弱者が淘汰され強者が残るという考えは当たり前のことだったはずだ。一方、淘汰自体の概念と比べると、あらゆる生物が子孫に伝えることができる個性を自然に発生させるという考えは、背景にあるメカニズムが全くわからないため、受け入れることは簡単でなかったはずだ。もちろん当時の人も、人間を真面目に観察しておれば、それぞれが個性を持っており、誰一人として同じでないことはわかっていたはずだ。しかしダーウィン進化論にとって個性とは、子孫に伝わる性質でなければならなかった。メンデル遺伝学が登場するのは十年後だ。それでも、種としての共通性を基盤として、生殖過程で自然に発生する変異の蓄積が個性を生み、それが子孫に受け継がれると考えたダーウィンの慧眼には瞠目せざるをえない。
次にこの19世紀の議論を、21世紀ゲノム時代の視点で見直してみよう。
今や何百もの種の全ゲノムが解読されている。ゲノムの大きさ、DNA複製のメカニズムを知れば、大腸菌でさえ一回分裂して2個の大腸菌になるとき、両方のゲノムが全く同じである可能性があるなどと考える人はいないはずだ。すなわち、分裂時、あるいは分裂後に個体ごとに異なる様々な変異がゲノムにおこり、ゲノムは多様化し、個性が生まれる。変異は、1代、2代と世代を重ねることにより蓄積し、個体数の増加とともに多様性は増大する。このとき、増殖を損なう変異がおこると個体の維持ができずに自然に淘汰される。逆に、増殖力が高まって他の個体の個性を凌駕することも可能だ。このように、原核生物の場合、生存と複製に必要なゲノム構造が損なわれない限り、種内のゲノム多様性が発生し続け、こうして生まれた個性が環境にフィットすると、今度はこの個性が優勢になり、新しい個性が種として共通の性質になる。
ゲノムのみが経験(自然選択)を記憶できる媒体である原核生物では、ゲノムの多様化はエントロピーの増加を意味する。こうして増加したエントロピーは、自然選択過程により環境にフィットした個性が選択されることで低下する。この過程が異なる環境で起こると、選ばれる個性は異なるため、それぞれの個性へと種全体が収束することで、環境に応じた種が誕生し、種の数は増加する(もちろん環境が同一だと、種は分化できない)。このエントロピーが増減を繰り返す過程の繰り返しが進化で、すべてのゲノムは38億年前の生命誕生まで遡ることができる。このように、ゲノムに蓄えられた過去の記憶はゲノムの個性に等しい。言い換えると、個性とは過去の経験の記憶により形成される多様性と言っていい。(図2)
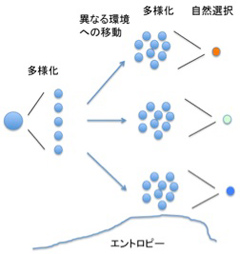
図2 個体のゲノムは自然に変異を蓄積するようにできている。これにより個性の多様化が起こる=エントロピーが増大する。環境に適応した個体が選択されると多様性は低下し、新しい性質が共通の性質になる。これが様々な環境で起こると、種は多様化する。
原核生物では、ゲノムの記憶がその生物の個性のすべてを決めていると言えるが、多細胞生物が生まれ、生殖細胞と体細胞が完全に分かれると、個体の個性は、ゲノムの個性とは言えなくなる。このことは私たち人間について考えるとよくわかる。
現在では同じ種でも個体間でゲノムが多様化していることがわり、確かにゲノムの個性が子孫に受け継がれ維持されていることは間違いがない。このゲノムの個性から、病気のリスクや、できれば他の様々な性質を予測したいと、個人遺伝子診断サービスが最近提供されている。しかし、個人の全ゲノムを解読したとしても、それですべての個性を予測できると考える人はいない。受精卵が発生を始めてから死ぬまで、我々は様々な経験を通して、ゲノムでは決まらない個性を作り上げていく。これは経験が記憶され、その記憶が個性となるからだが、この記憶はエピジェネティックスとして総称される。ただ、この時の記憶媒体は、クロマチン情報と神経ネットワークの両方(人間では言語も加わるが)からできている。例えば昔人気を博した一卵性双生児姉妹、金さん銀さんの性格や顔があれほど異なるのは、クロマチン情報の記憶と神経ネットワークの記憶の違いが原因だ。
まずクロマチン情報を介する個性について見てみよう。ゲノムの個性と比べた時クロマチン情報を介する個性を研究する難しさは、個体レベルの個性が一つの情報に代表されるのではなく、細胞一個一個の個性の集合として表現されている点だ。
まず発生過程で、何百種類もの細胞が分化するのは細胞ごとにクロマチン情報の違いが生まれるからだ。ただ、発生過程では、決まった細胞を繰り返し安定に分化させることが要求され、クロマチン情報が間違って多様化しないように設計されている。もちろん、この過程でも間違いは起こり、その間違いが細胞の生存や機能を損なわない限り、細胞の記憶として維持される。以前述べたように、クロマチン情報も細胞分裂を超えて維持することができるため、発生の初期に起こった間違いは、多くの子孫細胞に受け継がれる。このような間違いは、時間を経て問題になる場合がある。
有名な例は、1944年の冬、ナチスドイツに封鎖され極度の飢えに苦しんだアムステルダムで胎児期に飢えを経験して生まれた人たちは、高齢になってからインシュリンの分泌が低下し糖尿病になる。これは、胎児発生時に飢えにさらされて起こったクロマチン情報の間違いが、高齢になってから細胞に影響し始めたことを示唆している。同じように、胎児期にお母さんがアルコールを摂取すると、様々な細胞でクロマチン情報の記憶間違いが起こることが知られている。
幸い、クロマチン情報の記憶形成は間違いが少ないように設計されており(増殖因子や分化因子がきめ細かく細胞の方向性を調節する)、それぞれの細胞でおこる記憶間違いはランダムに起こるが細胞の生存から見て許容範囲内で止まっている。またクロマチン情報の変異は細胞ごとの個性になるため、変異が許容範囲を超える細胞は淘汰されるが、個体全体には影響が少ないことが多い。
同じ臓器で同じ細胞が作り続けられるためにはクロマチン情報は必須で、それを維持することは重要だが、発生後も外界のストレスによりクロマチン情報は少しづつ変化する場合が多い。例えば、老化によりクロマチン情報が変化することが知られている。しかし、このような変化は個々の細胞レベルで止まり、各細胞で全くランダムに起こるため、臓器や個体の機能にとって問題にならない。しかし、この個々の細胞にランダムに蓄積した様々なクロマチン情報の変化が集合したものが、クロマチン情報を介する個性として表現される。
以上をまとめると、クロマチン情報の記憶は、各細胞個別に形成される。この記憶は細胞の種類に応じて同一になるよう設計されているが、どうしても小さな記憶違いが生じて、これが細胞の個性を形成する。そして、この細胞の個性が集まって、個体の個性になるが、ゲノムの個性と異なり、個体レベルのクロマチン情報の個性を一つの情報で代表させることはできない。また、クロマチン情報を介する記憶や個性は、各細胞に限局されており、統合されることはない。個体レベルのクロマチン情報の個性は、あくまでも細胞の個性がただ集まっただけの結果だ。
エピジェネティックスな変化の中には、神経ネットワークを介する情報の記憶も当然含まれる。事実、私たち人間の個性は、脳の個性として表現される部分が多い。例えば、生後訓練を繰り返して筋肉が発達し、早く走れるようになる過程にアメフラシで見たのと同じ神経ネットワークを介する記憶が大きく寄与している。もちろん、筋肉自体も発達し、これにはクロマチン情報の変化も寄与するが、神経系の記憶が存在しないと筋肉は発達しない。残念ながら、こうして訓練した結果は、次の世代に受け継がれず、一代限りだ。
神経ネットワークのメカニズムを突き詰めると、細胞が発生し、成長し、老化する過程で起こるクロマチン情報の記憶と同じと言っていい。どんなに複雑な脳神経系でも、進化過程でゲノムに蓄積された記憶情報に基づいて空間的・時間的に発現する特異的なシグナルに従って順々にクロマチン情報が変化した細胞を分化させ、これをゲノムの記憶に従って立体的に構造化させることで、形成される。また、生前、あるいは生後それぞれの神経が受ける刺激に対しては、一次的な反応が細胞内シグナル伝達経路の活動として反応する。そしてシグナルの質や強さに応じて、前回見たように、神経細胞のクロマチン情報が変化し、長期記憶が成立する。しかし図3に示すように、神経細胞の反応は他の体細胞の反応と全く変わるところはない(図3)。
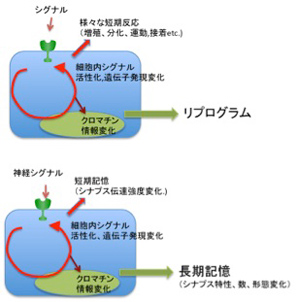
図3:神経細胞と一般体細胞の比較。
説明は文中。
一方、一般の体細胞と神経細胞の記憶は、個々の細胞内で起こる変化が個体レベルで統合されるかどうかという一点で大きく異なっている。すでに述べたように、クロマチン情報の記憶に起因する細胞の個性は、ただ集合して個体の個性に寄与するだけだが、神経系では細胞レベルのクロマチン情報の変化に起因する個性は、細胞同士がネットワークを形成することで、個体レベルで統合される。この統合は、神経細胞にとどまらず、筋肉やホルモン産生細胞など、多くの体細胞をも統合できる。
実際には、同じような細胞の変化を統合して個体の個性を変化させることは、内分泌システムにも見ることができる。オタマジャクシがカエルへと変態を遂げる時、甲状腺ホルモンが分泌され、体全体の細胞を統合する。同じことは、私たちが思春期に急速に様々な性的特徴を発生させるのも同じだ。しかし、一見個体全体を統合している高等動物の内分泌システムを詳しく見てみると、ほとんどがどこかで神経系とつながっており、神経ネットワークの記憶の支配を受けていることに気づく。
このように、神経細胞は、神経細胞にとどまらず様々な細胞と直接、間接にネットワークを形成する能力を開発し、全く新しいレベルの個体性の統合を可能にしている。
これに加えて、神経ネットワークでは、視覚、聴覚、嗅覚など、外界の変化を迅速に、立体的に捉え、この入力を個体として統合された神経ネットワークにリンクさせることができる。これにより、環境を全く異なるレベルで自己化することが可能になった(例えば景色の記憶は、環境を自己化したと言える)。もちろん、外界の自己化は原始ゲノム発生以来、生物進化の駆動力と働いてきたことだが、神経系による自己化は、あるがままの外界を表象し自己の延長として自己化する点で、これまでの自己化とは全く異なる。
この個体内外の統合と、自己の発生についてはわかりにくいと思うので、次回は、「統合と自己」というタイトルで、神経系ネットワークが可能にする個体と細胞、個体と外界の統合能力について解説してみたいと思っている。