

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
脳の自己
2017年2月15日
脳を頂点とする階層的な神経回路による新しい情報システムの誕生で可能になったことについて書いた前回の内容をまとめると、
- 1)様々な物理的刺激を受容することが可能になり、個体が経験する環境の範囲が急速に拡大する。
- 2)結果、生物はほぼ無限とも言える刻々(ms単位で)変化する情報にアクセスし、またそれに対する迅速な反応が可能になった。
- 3)神経回路に対する体内・体外からの様々な刺激を脳内の神経回路内に表象し、そのイメージを記憶することが可能になった。
になる。
古来人間の最大関心事である自己や意識はこれを基盤として新たに生まれた高次情報だが、その発生のためには、脊椎動物の脳構造の大きな変化が必要だったので、まず構造の進化を簡単に確認しておこう。
脳の急速な増大
脳神経回路を媒体とする新しい情報システムの進化過程では、一貫して脳の神経細胞数が増え続けてきた。例えばマウスは約2億個のニューロンがあるが、人間ではその数は1600億個にのぼる。闇雲に回路を複雑にすれば高次機能が生まれるわけではないが、素子数を増やすことで発展してきたコンピュータを見れば、まず細胞数を増やすことが必要であることは直感できる。実際全く異なる情報システムであるゲノムも進化の過程で塩基数が増大し、ゲノム内の構造も複雑化してきた。
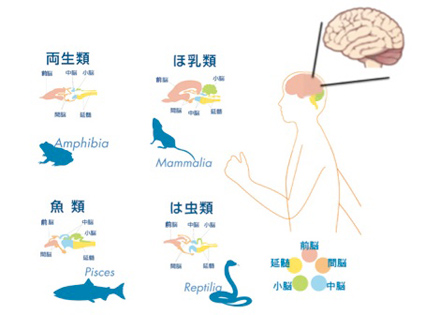
図1:進化の過程で前脳が急速に拡大する。これに続いて、サルやヒトでは皮質を陥入させ脳のシワ(脳溝)を形成することで、体積以上の皮質の拡大が可能になった
脳は大きく分けて、後脳、中脳、前脳、および嗅球に分けることができるが、図に示したように脊椎動物進化では、もっぱら前脳を拡大させてきた(図1)。
前脳皮質の神経細胞を増加させるメカニズムだが、脳室のsubventricular zone (SVZ)で水平に幹細胞を未分化なまま増殖させて皮質面積を拡大するとともに、増殖している幹細胞が今度はradial glial cellに沿って縦に細胞を供給し、順番に異なるタイプの神経細胞へと分化することで縦に厚みが増すことで達成されている(図2 Montiel et al, Journal of Comparative Neurology, 524:630, 2016 より転載)。
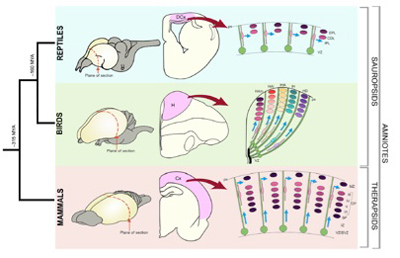
図2 哺乳動物、鳥類の皮質神経細胞を縦に並べていく神経形成の様式について示している。神経細胞を作る幹細胞がまず水平に広がり、この細胞から造られた神経細胞はradial glial cellに沿って移動しながら分化し、細胞数を増やす。
ただ脳の表面をそのまま増大させるだけでは、脳のサイズという物理的限界に当たる。そこで脳進化の過程で、皮質が立体的に陥入するいわゆる脳のシワ(脳溝)が形成される様になり、このおかげで、サルから人間の進化の過程で脳の皮質面積は急速に増大する(脳溝は図1の人間の脳参照)。
同じ幹細胞に由来する異なるタイプの神経細胞が縦に並ぶことで脳皮質の層構造(6層)ができる。この縦に並んだ細胞からできる層構造が最小セットとなり、この単位が横に束ねられ、細胞同士がネットワークを作ると、カラムと呼ばれる脳の最小機能単位が出来上がる。このカラム構造では、各層の神経細胞の反応性は似通っている(例えば同じ方向の動きに反応するなど)。この様に、脳では細胞数がただ増えるのではなく、小さな単位のネットワークが集まってモジュールができ、それがまた集まってという様に、ネットワークは階層的に拡大していく。
以上が脳のサイズ拡大を支える解剖学・細胞学的の知識の基本だ。この基盤の上に、自己や意識が生まれる。
自己の誕生
自分の行動を他の動物と比べ、また自分の過去を振り返れば、自己や意識が脳の進化とともに現れ、また「物心つく」と言うように、自己や意識が一人一人生まれてから新たに発生することがわかる。もちろん古来、自己や意識は人類にとっての最大の謎で、哲学から生物科学、神経生物学、心理学まで多岐にわたる議論が続けられてきた。当然この話題についてのすぐれた著作は多く、本当は私の出る幕ではない。しかし脳神経回路を媒体にする情報の理解なしに、生物の情報の進化を構想することはできないので、この問題についても私なりの理解をまとめておくことにした。
まず問題は、自己から始めるか、意識から始めるか、あるいは同時に扱うかだが、両者は脳の最も高次の機能として密接に関連し合っているため、どちらがより重要とか、どちらが高次とか決めることはできない。
脳神経系以外の自己
そこで「エイヤ!」と「自己の誕生」から始めることにした。というのも、自己という言葉は、脳神経回路情報だけでなく、様々なレベルの生物活動に対して使われる。私自身も原始ゲノム誕生について説明した時、「ゲノムの自己性」と、この言葉を用いた。実際、私たちは「細胞の自己」のように脳神経系が存在しない生物に自己を使っている。例えば自己複製という言葉は最もポピュラーだろう。他にも、昨年のノーベル賞に輝いた大隅良典さんの「オートファジー」も、細胞の自己を想定した使い方だ。
これらの場合で、「自己」の意味する内容を考えてみよう。ゲノムの自己と私が呼んだ場合はひと塊りの核酸が周りから完全に分離し、その上で自律的な複製能力を持つことが条件となっている。これが単細胞動物となると、内と外を隔てる細胞膜が存在するため周りからの分離はさらに明確になる。ただ、シャボン玉のように膜で周りから分離される状態が維持できても、自己性があるとは思わない。周りの環境から完全に分離するとともに(自己と外とに境界線を引くことができる)、同じ細胞(境界の内側)を自律的に複製できる能力があって初めて自己性があると言える (熱力学的条件などは自己複製で全て表現できている)。周りから完全に分離して、自己複製能を持つことが、「自己性」成立の条件だ。
多細胞動物になっても、個体の内と外ははっきりと分けることができるため、細胞と同じ意味で「個体の自己」を使うことができる。ただ場合によって「個体の自己」と「細胞の自己」が分離することがある。最も分かりやすいのがガン細胞で、個体から独立して勝手に増殖すると同時に、ゲノムも個体とは異なっていると言える。この極端な例が、ハマグリやタスマニア・デビルに見られる、個体から個体へと伝搬するガン細胞だろう。このようなガン細胞は自己性が高いと言えるかもしれない。
他に「自己」が定義されている分野が免疫学だ。
免疫学はジェンナーによる種痘の成功により認識された「2度なし現象」(一度感染すると次に感染しない)についての研究がはじまりで、パストゥールの狂犬病ワクチン(抗原の概念)、そして北里・ベーリングによる免疫血清療法(抗体の概念)の開発により、外来抗原に対する特異的な抗体反応という概念が確立する。
このように最初、外部から侵入する病原体への防御反応機構として始まった免疫学は、ランドシュタイナーらによる血液型の発見により、免疫系が自己の成分と非自己の成分を区別できることが明らかになる。その後、臓器や組織の移植研究から、移植抗原が(MHC)が異なる他人からの組織に対してだけ強い免疫反応が誘導されることが明らかになると、「自己と非自己の区別」は免疫学の最重要課題になる。
20世紀後半に始まる免疫学の大きな進展により、免疫系が自己成分とそれ以外をどう区別するかについてはすでに詳しくわかっている。詳細を省いてこのメカニズムを見てみよう。
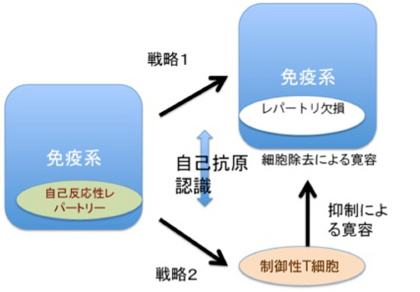
図3:免疫系の自己と非自己の識別:説明文中
免疫反応にはT細胞とB細胞が関わっているが、原則的にはどちらの細胞も自己の成分に対しては反応しない。これを、自己成分に対する免疫寛容と称している。この寛容には2種類の戦略が存在している。最初の戦略は、リンパ球の分化がまだ未熟な段階で抗原刺激を受けると、成熟後におこる細胞増殖や分化の代わりに細胞死が誘導される。発生初期から体内に存在する自己抗原は、未熟な段階のリンパ球と反応することができこの結果自己の抗原に反応するリンパ球の細胞死を誘導して免疫システムから除去している(図3)。この過程は、T細胞では胸腺内、B細胞は骨髄内で行われる。
もう一つは我が国の坂口志文らにより明らかにされた戦略で、細胞死による寛容とは逆の戦略と言える(図3)。この戦略は制御性T細胞と呼ばれる特別なポピュレーションだけで見られる。制御性T細胞が自己抗原と出会うと、細胞死の代わりに活性化され、長期間体内で生存できるようになる。これにより後にキラーT細胞のような他のリンパ球集団が同じ自己抗原に反応しようとするとき、その細胞を抑制し、自己抗原に対する反応を抑制する。
このように免疫系の多様な認識システムは、発生過程で生成的に形成される。こうして生成される免疫系の自己とは、免疫系の抗原受容体が認識できる自己抗原の総体と定義でき、これらの抗原は免疫系が反応できないレパートリーとして免疫系に表象される。言い換えると、自己抗原は発生初期の選択の結果、細胞死による「レパートリー欠損」として、あるいは自己反応性抑制性T細胞レパートリーの長期維持として表象されている。自己の範囲に境界線を引くとすると、免疫の自己とは、成熟後の免疫系が反応できない抗原として線引きができる。
実は免疫システムにはもう一つの自己認識がある。それはMHC restriction(組織適合抗原拘束性)と呼ばれるT細胞の認識方法だ。抗体と違ってT細胞の抗原受容体は、抗原をそのまま認識することまずない。代わりに、図4に示すように、まずタンパク質はプロテアソーム(タンパク質分解酵素複合体)でペプチドへと分解され、それが自己のMHCと結合したときに新たに生まれる分子構造を認識している。
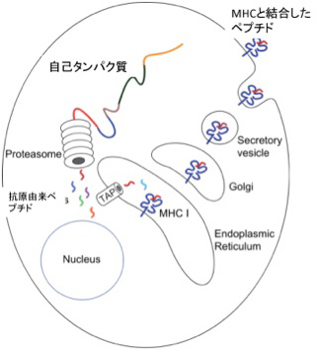
図4 T細胞抗原受容体は、一度自己MHCに参照できたペプチドだけを認識する。Wikipediaを改変転載。
言い換えると、自己、非自己を問わず、T細胞抗原受容体に対する抗原は小さなペプチド+自己MHCで、抗原は自己のMHCと結合するかふるいにかけられた後、T細胞抗原受容体で認識できるようになる。すなわち、抗原の段階でまず自己に参照される。ただこの場合も、免疫側にとって自己ペプチド+自己MHCを含む、T細胞抗原受容体が反応できない全ての抗原が自己と認識され、これは抗原に反応できるT細胞受容体のレパートリー中にぽっかりと空いた欠損として表象される。
以上、脳神経系の自己に行き着く前に、脳神経系とは異なる2種類の自己の定義について見てきた。この2つの自己をまとめておくと、
- 1)外界との境界を設け、境界内を複製できることで生まれる自己、
- 2)生成的に形成される認識系に区別して表象されることにより定義される自己
の2種類があることがわかってもらえたと思う。
一見これらは、脳神経系の自己とは接点がないように思えるかもしれないが、脳神経系に生まれる自己性は、この2種類の自己と多くの共通性を持っている。次回はいよいよ、脳神経系に生まれた自己について見てみよう。