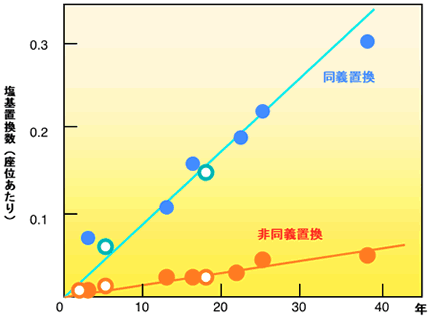
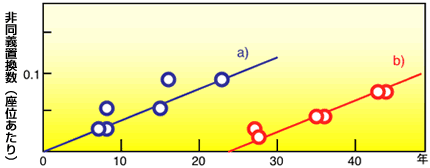
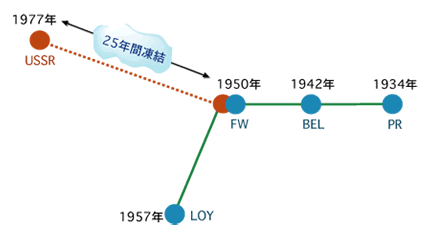
最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。 バックナンバー 【25年の眠りから覚めたインフルエンザウィルス:ウィルスの分子進化学】 2005年8月12日 平成17年4月14日付けの毎日新聞に身の毛がよだつような記事が載っていた。1957年から1958年にかけて、全世界で400万人の死者を出したといわれている、通称「アジア風邪」が大流行した。このインフルエンザウィルスは「H2N2」型だが、この型のインフルエンザは1968年以降起きていない。記事によると、アメリカの医療会社がこのウィルスのサンプルを世界中の6500施設に誤って送付してしまったらしい。幸い、このウィルスが漏れて人に感染したという報告はまだないようだが、漏れでもしたら、1968年以降に生まれた人は免疫を持たないので、若い人を中心に大流行する可能性があった。なんとも物騒な話である。 今から20年ほど前に筆者のグループは、上記と同じ心配な事態が実際に起きていたらしいことを、インフルエンザウィルスの分子進化学的研究からつきとめ、論文に報告している。この論文で筆者らは、ウィルスにも普通の生物と同様の分子進化学があることを発見した。その分子進化学を基礎にすると、おかしな振る舞いをするデータが見つかったわけである。いささか古いデータだが、その顛末も含めてここで紹介してみたい。インフルエンザの流行のしくみインフルエンザウィルスの分子時計恐怖の復活 インフルエンザの流行のしくみ よく知られているように、インフルエンザはインフルエンザウィルスと呼ばれるウィルスの一種によって引き起こされる。この風邪の流行はすでにギリシャ・ローマ時代にも知られていたようで、その報告が今でも残っているらしい。インフルエンザは感染の規模が大きいことで有名である。1918年ー1919年にかけて世界的に大流行したインフルエンザ(通称スペイン風邪)は規模の大きさで群を抜いている。世界中の死者の数は1500万人から2500万人にも上ったと推定されている。これは人類がこれまで経験した最大の災禍だといわれている。人類はさまざまな疫病を克服してきたが、インフルエンザは、エイズと共に、人類に残された最大の疫病といっても過言ではなかろう。 世界規模で起こる疫病だからこそ、その対策も全世界的に行われている。世界のどこかでインフルエンザが発生すると、世界保健機構に報告され、そのウィルス株は単離され、凍結保存されている。こうしてストックされたウィルスサンプルはワクチンの開発やウィルスの研究に利用される。 インフルエンザウィルスは血清学的に A, B, C の3つの型に分類され、A 型はしばしば大流行を起こす。2本鎖DNAを持つ通常の生物と違って、インフルエンザウィルスの染色体は一本鎖RNAでできている。通常、RNAを染色体に持つRNAウィルスでは、一本の長いRNAに幾つかの遺伝子を乗せているのだが、面白いことにこのウィルスでは、8本のRNAからなる分節構造をとり、各分節は1つないし2つの遺伝子を暗号化している。この独特な分節構造がインフルエンザの大流行に関係がある。 インフルエンザウィルスはほぼ球形をしていて、その表面には、赤血球を凝集させる働きがある赤血球凝集素ヘマグルチニン(通常HAと書く約束になっている)と、酵素の働きを持つノイラミニダーゼ(NA)とがある。HAとNAは宿主の抗体によって抗原として認識される。従って、これらの2つの分子が突然変異を受けて変化すると、抗体は認識できなくなり、逆にウィルスは抗体からの攻撃を逃れて蔓延することができるわけである。ウィルスの内部にはそれ以外のタンパク質と8本のRNAが詰まっている。 普通、ある特定のウィルスは宿主とする生物種の種類が狭い範囲内に限定されているのだが、困ったことに、ヒトのインフルエンザウィルスはヒト以外に、家畜や鳥にも感染できる。家畜や鳥に感染したヒトのウィルスはときどき家畜や鳥のウィルスのRNAと一部を取り替え、再びヒトの集団に感染することがある。取り替えたものがHAやNAだと、いわば新しい着物を着たようなもので、それに対する抗体が人間の集団にまったくないので、ウィルスは大流行する。ウィルスのRNAが文節構造を取っていることが、遺伝子の取り替えを容易にしている。大流行と大流行の間に小流行があるが、これはHAやNAのアミノ酸が別のアミノ酸に置き換わった結果起きた変異に由来する。小流行では世界規模でインフルエンザが蔓延することはなく、せいぜい地域的流行にとどまる。 HAやNAには幾つかの異なるタイプがある。HAとNAの組み合わせで色々な型、すなわち亜型ができる。これまで知られているヒトのインフルエンザウィルスの亜型は3つあり、H1N1,H2N2,H3N2と略記されている。ところが鳥にはたくさんの亜型があり、ヒトの3つの亜型すべては鳥の系統の亜型に見つかっている。おそらくヒトの集団に現れるインフルエンザウィルスの亜型の源流は鳥のウィルスにあり、鳥のウィルスにある亜型をときどき引っぱり出してきては、ヒトに大流行を引き起こすのであろう。近頃ニワトリのインフルエンザがしきりと話題になっているが、その背景にはこうした事情がある。 ▲このページのトップへもどる インフルエンザウィルスの分子時計 インフルエンザウィルスも普通の生物と同じように進化するのだろうか。そして宿主の遺伝子にみられたように、ウィルスの遺伝子も分子進化の中立説に従うのであろうか。こうした疑問に答えるべく、林田秀宜(現、奈良県立医科大学教授)を中心とする筆者のグループは、それまでに決定されていたA型インフルエンザウィルスの塩基配列を用いて1983年頃から研究をスタートした。 インフルエンザが流行するたびに、そのウィルス株が凍結保存されるので、それはいわばウィルスの化石のようなものとみなせる。しかも完全な遺伝情報を手にすることができる化石である。また、すでに述べたように、HAとNAの組み合わせで色々な型、すなわち亜型に分類される。結果的にわかったことだが、HAとNAの組で決まる亜型は生物種のようなもので、同系(オーソログ)か異系(パラログ)がおおよそ判断できる。 このことを利用するとインフルエンザウィルスに分子時計が存在するかどうかを確かめることができる。例えば、同じ亜型のウィルスの一つの遺伝子について、T1年に流行したウィルスのサンプルの塩基配列とT2年に流行したウィルスのサンプルの塩基配列を比較して、塩基が異なっている座位の数からT12(≡T2 −T1)年の間に貯めた座位あたりの塩基置換数K12を推定する。幾つかのサンプルの組から、K - Tの関係、すなわち分子時計を検証できる。 林田らは、3つの遺伝子、すなわち3つの分節でK - Tプロットを調べたが、いずれにもみごとな直線関係、すなわち分子時計の存在が確認された。図1に分節8の結果を示した。さらに同義置換の速度(直線の傾きで示される)は非同義置換の速度より大きい。ここで、同義置換とは、タンパク質を暗号化している遺伝子領域で、一つの塩基を別の塩基で置き換えたときに、アミノ酸が換わらない場合である。変わる場合を非同義置換という。前者は塩基を変えてもタンパク質に変化が起きないので、この変化は機能的制約が極めて弱い部分の変化とみてよい(本シリーズ:「分子の進化速度は形態と分子の進化を橋渡しできるか」を参照)。図1は、機能的制約が弱く働いている同義置換は制約が強く働いている非同義置換より進化速度が大きいことを示している。この結果は、分子時計の存在と共に、ウィルス遺伝子の進化が分子進化の中立説で説明できることを意味している。さらに、同義置換速度は異なる遺伝子間でほぼ等しいが、非同義置換速度は遺伝子ごとに大きく異なる。この性質も普通の生物の進化のパターンと全く同じであり、中立説の枠内で理解できる。 図1.インフルエンザウィルスの分子時計 普通の生物との類似点はこれまでで、ウィルスの進化の速度は桁外れに大きい。ウィルスの同義置換速度は0.01座位/年で、これは哺乳類の遺伝子のおよそ数百万倍にものぼる。通常の生物の分子の進化は数百万年、すなわち、地質年代を単位として起こるのに比べ、ウィルスの分子は年を単位として起こるわけである。インフルエンザウィルスはまさに進化のミニチュアと呼ばれるにふさわしい。 インフルエンザウィルスは大きくA型、B型、C型の3つの型に分類されるが、この順に流行の程度が小さくなる。それと対応して進化速度も順に小さくなる。このことは突然変異の要因が一様に降り注ぐ放射線のようなものではなく、染色体DNAや染色体RNAの複製に際して生じる複製エラーであることを示唆している。 インフルエンザウィルス以外のウィルスについても同じように進化速度を求めることができる。エイズのウィルスをはじめとして、色々なウィルスの進化速度を、宿主の遺伝子の進化速度と比べてみると、いずれのウィルスも極めて速いスピードで進化していることが分かる。ことに、エイズウィルスはインフルエンザウィルスより進化速度が大きい。 こうして、インフルエンザウィルスの進化速度が定量されたが、宿主の遺伝子の数百万倍にも及ぶ高い変異性を持つことが明らかになった。このウィルスでは地質年代(100万年)の単位で起こる通常の生物の進化現象がわずか数年の単位で起こる。この高い変異性がワクチンを作ることを困難にしている理由であろう。HAやNAがどんどん変わるので、せっかく作ったワクチンも、直ぐに役立たずになってしまう。 ▲このページのトップへもどる 恐怖の復活 この解析の途中で、著者のグループは、インフルエンザウィルスの進化には奇妙な振舞があることに気がついた。すなわち、ある時期にいったん進化を停止し、何年か後に再開することがあるのだ。1983年筆者のところで、大学院生の林田君の指導で、卒業研究をしていた4回生の藤博幸君(現、九州大学大学院教授)はN1亜型のインフルエンザウィルスの分子時計を調べていた。ある日、解析結果をもって林田君と一緒に私の部屋へやってきた。まず、4つのウィルス株(PR、BEL、FW、LOYと略記する)の間で置換数Kと経過時間Tの間の関係を私に示した。KとTの間にはみごとな直線関係があった(図2a)。もう一枚、USSR株をPR、BEL、FWと比べた図を恐る恐る出して私に見せた(図2b)。その図を見て私はびっくりした。この図は直線関係を示していたが原点を通っていなかった。私はいきなり藤君に、「経過時間がゼロなら置換数もゼロのはず。だからK-T図は原点を通らなければいけない。基本的なことだ」と叱った。するとそばにいた林田君が「でもどうしてもこうなるのです」と助太刀した。その場でもう一度詳しく調べ直して、この図が正しいことが判明した。 図2.NA遺伝子の分子時計 この図2bは傾きも図2aと同じで(すなわち進化速度が等しい)、明らかに分子時計の性質を示している。しかし、25年分だけ右にシフトしている奇妙な図になっている。これは明らかに1977年に流行したUSSR株(通称、ロシア風邪)に原因がある。文献を調べてみると、この株の塩基配列は不思議なことに1950年に流行した株に非常によく似ていることが報告されていた。 この奇妙な現象の最も合理的な解釈は、USSR株が1950年と1977年の間の25年間だけ進化を完全に停止し、その後再び同じ進化速度で進化を再開したということである(図3)。なぜこんな奇妙なことが起こるのであろうか?考え易い説明の一つは、どこかで凍結保存されていたUSSR株が、何らかの理由で1977年に外に漏れたということであろう。 図3.インフルエンザウィルスN1サブタイプの系統関係と進化の凍結 もしそうなら、ことは重大である。そう直感した筆者らは、確かな証拠が出るまではこの現象を「進化の凍結」という主張にとどめることにした。1983年の学会で筆者らは以上の成果を報告したが、その際「サンプルが外に漏れたのではないか?」という質問を大変恐れ、あらかじめ答えを用意しておいた。実際、そういう質問があった。私の答えは記憶している限り、「風が吹けば桶屋が儲かる」式のものであった。すなわち、1950年にシベリアでFW株に感染して死んだ人あるいはトリが凍結し、25年後にたまさか氷解したところをトリがついばみ、それが人に感染した。本人も含め、会場にいたほとんどの人はこの説明に納得していなかったことはいうまでもない。 理由はともあれ、一時期進化を完全に停止し、25年もの後に進化を再開するということがあれば、その間に生まれた若い世代はこのウィルスに対する抗体をまったく持たないので、インフルエンザは若い世代を中心に大流行するはずである。インフルエンザの大流行はこうした理由でも起こりうるということだが、人間の不注意がもたらす災害の中で、これほど大きな災害はあるだろうか。 文献:Hayashida,H., Toh,H., Kikuno, R. and Miyata,T. Evolution of Influenza Virus Genes. Mol. Biol. Evol. 2, 289-303 (1985) ▲このページのトップへもどる [宮田 隆] 宮田 隆の進化の話 最新号へ