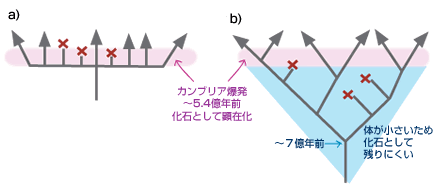
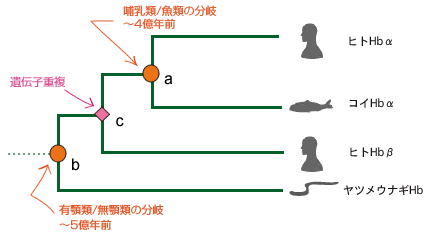
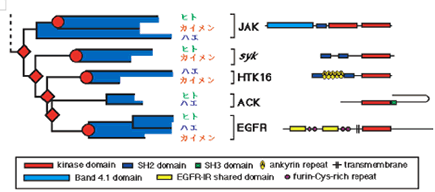
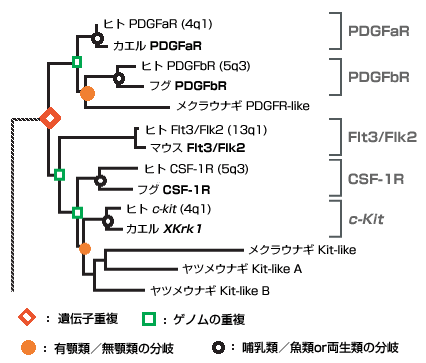
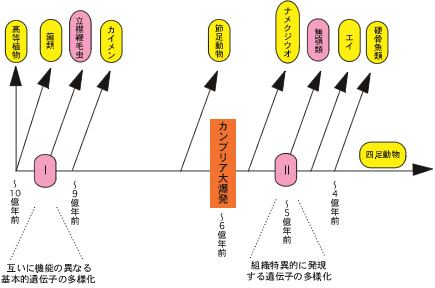
最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。 バックナンバー 【古い遺伝子を使って新しい形を作る:カンブリア爆発と遺伝子の多様性】 2006年3月1日 およそ5億4千万年前、カンブリア紀と先カンブリア時代の境で、生物の全歴史を通じて特筆すべき事件が起きた。その規模においでも、デザインの斬新さにおいても、どの時代の生物にもひけを取らない、さまざまな多細胞動物が爆発的に出現した。これは生物進化史上最大のイベントで、カンブリア爆発といわれている。この爆発的多様化は、新しい遺伝子を作ることなく、すでに単細胞の時代に作ってあった遺伝子を再利用して起きたらしい。問題をほぐすカンブリア爆発カンブリア爆発の規模と原因遺伝子重複が起きた時期を推定する遺伝子族:サブファミリーと組織特異的遺伝子遺伝子族の多様化とカンブリア爆発との関連立襟鞭毛虫:単細胞原生生物に多細胞用の遺伝子セット遺伝システムの柔軟性とソフトモデル形態進化の便宜主義と遺伝子進化のソフトモデル 問題をほぐす 本シリーズの第一回でカンブリア爆発と遺伝子の多様性の問題を取り上げたが、短く書かれたもので、ほぼ結論のみの記述で終わっている。カンブリア爆発は重要なテーマであるので、最近の発展も加味しながら再度論じてみたい。 ヒトゲノムの全塩基配列が決定された際、日本の主催者側から報道関係者に説明があったようである。その折、ヒトの全遺伝子数は約37000でショウジョウバエのおよそ2倍にあたるとの報告があったらしい。それを聞いた報道関係者の一人が私の研究室にやってきてこうつぶやいた。「ヒトの全遺伝子数がショウジョウバエのたかだか2倍とは驚いた。複雑さから考えて、とても信じられない。」私は即座に、「全然驚かない」と答えた。そのわけは以下を読んでいただければ分かるであろう。 我が国が世界に誇る集団遺伝学者で「分子進化の中立説」の提唱者である木村資生(本シリーズ、「パラダイムシフト:分子進化の中立説」を参照)は、彼の晩年の著書「生物進化を考える」(1998年、岩波書店)で次のように述べている。「今後に残された大きな問題の一つは、表現形レベルの進化と分子レベルの進化との間にどうしたら橋渡しができるかということである。この方面でも、将来、日本の若い研究者によって世界に誇ることができるような業績が上げられることを望みたい。」 この問題は、別の言い方をすれば、生物の進化と遺伝子の進化の関連を問う問題であり、生物進化の分子機構の問題である。さらにこのテーマは、分子進化学誕生当初からの大問題でもある。ただ、この問い方ではあまりにも漠然としているので、具体的に研究を始めるにあたって問題を解きほぐす必要がある。 生物が示す種の多様性は生物の最も大きな特徴の一つである。「生物の進化と遺伝子の進化の関連」という問題を、「生物の多様性と遺伝子の多様性の関連」としてみよう。さらに生物の多様性に対して、下で詳しく述べるが、多様な形態を持った多細胞動物が爆発的に出現したカンブリア爆発という具体的な“事件”を当てはめてみる。そしてこう問うてみよう:カンブリア爆発が起きたとき、遺伝子も爆発的に多様化したか? かなり具体的な問題になったが、さらに“遺伝子”に対しても限定しておこう。 いうまでもなく、多細胞動物の体は複数の細胞で構成されている。細胞の一つひとつには共通のDNAがあり、DNAには数万の遺伝子が乗っている。これらの遺伝子にはバクテリアも含めてすべての生物に共通に存在する遺伝子もあれば、多細胞動物だけが持つ遺伝子もある。例えば異なる細胞同士がコミュニケーションしたり、動物の体を作るために必須な遺伝子などがそれにあたる。ここでは多細胞動物に特有の遺伝子を問題にするのが適当であろう。そうすると問題は以下のようにほぐされ、具体的に扱うことが可能になる。すなわち、「カンブリア爆発に伴って、多細胞動物に特有の細胞間情報伝達や形態形成に関与する遺伝子も、爆発的に作られたのであろうか?」この問題は現在の技術で十分アプローチ可能である。 さて、遺伝子の多様化機構はすでによく理解されている。タンパク質は、一度獲得した機能を長い進化の過程で保持し続ける。ここからは新しい機能を持った遺伝子は進化しない。どうやって新しい機能を持った遺伝子が進化するのか?生物は細胞内の体制が出来上がった太古の時代から、一つの巧妙な方法で膨大な数の遺伝子を生み出してきた。その方法は、既存の遺伝子の機能を保持したまま、機能的革新をもたらすものでなければならない。生物が採用した方法は遺伝子のコピーを作ることであった。このことを遺伝子重複という(本シリーズ「制約からの解放と革新への道」を参照)。 もう一つ重要な遺伝子多様化の機構が知られている。コピーされたいくつかの遺伝子あるいはその一部を組み合わせて、一つの大きな遺伝子に統合することで、新しい機能を持った遺伝子が作られる。このことを遺伝子混成(ジーン・シャフリング)という。遺伝子混成は、わずかな数の遺伝子から多様な遺伝子を創出することを可能にする。真核生物は遺伝子重複と遺伝子混成の機構で、驚くほど多様な遺伝子を生み出してきた。コピーとそれが基本となった組み合わせこそ、遺伝子多様化の基本的メカニズムなのだ。 進化の過程で遺伝子重複が繰り返し起こると、よく似た配列を持つ遺伝子のファミリー、すなわち遺伝子族が形成される。なかには匂い分子のレセプター遺伝子のように、数百のメンバーから形成されている遺伝子族もある。 ここまでくると問題は単純になる。問題は、「多細胞動物に特有の細胞間情報伝達や形態形成に関与する遺伝子の遺伝子重複がいつ起きたか?」という具合に、具体的に扱うことが可能な形に変形された。遺伝子重複が起きた時期の推定は、後で詳しく述べるが、分子系統学的な手法で可能なのだ。 ▲このページのトップへもどる カンブリア爆発 多細胞動物は、分類学上、動物界に分類されていて、最も原始的なカイメンや、より複雑な体制を持ったクラゲやイソギンチャクなどの二胚葉性の動物、さらには脊椎動物や節足動物など、多数の種類の動物を含む三胚葉性の動物が含まれる。動物界の最大の分類単位は門で、異なる門の間では体の形が全く異なっている。カンブリア爆発によって出現した動物は現在の門に相当する動物たちの祖先なのだ。彼らはすべて数百万年ほどの、地質学的には短い期間に爆発的に出現したといわれている。 カナダの西部、ブリティッシュ・コロンビアにバージェス頁岩と呼ばれる有名な化石産地がある。ここでは、カンブリア爆発当時の動物たちの姿が、まるで現実に見ているように、見事に保存されている。米国の古生物学者チャールズ・ドリトル・ウオルコットによって1909年に発見された。それに続いて、英国の3人の古生物学者、ハリー・ウィッテトン、デレク・ブリッグス、サイモン・コンウェイ・モリスはバージェス頁岩の重要性を現在の地位にまで高めることに成功した。今やバージェス頁岩は古生物学の聖地といったところで、古生物学者ならだれでも一度は訪れたか、あるいは行きたいと望むことであろう。 バージェス動物群の化石はカナダだけではなく、中国の雲南省の澄紅(チェンジャン)、グリーンランド北部のシリウスパセットでも発見されている。こうした事実は、バージェス動物群は限られた地域に生息していた動物ではなく、カンブリア紀には全世界的に繁栄していた動物だったということを示している。 カンブリア爆発は古くから生物学者の関心を引いてきた。ダーウィンも強い関心を持った一人で、自然選択の力で徐々に生物が進化すると考えていたダーウィンは、カンブリア爆発を深刻に受けとめ、現時点では説明不能とするしかないと認めている。一方、生物の創造神話を信じる人たちからみれば、カンブリア爆発は謎でもなんでもなく、まさに神が生物を創造した瞬間だったのだ。もちろん現在ではダーウィンが正しかったことを化石の証拠が示している:多細胞動物と思われる化石が、カンブリア紀よりずっと古い、9億年前の地層から見つかっている。また、20億年前に棲息していた単細胞真核生物の化石もあれば、35億年前に生きたバクテリアと思われる痕跡さえある。今ではカンブリア紀と先カンブリア時代の境で突如生物が誕生したのではないことは確かだ。 ▲このページのトップへもどる カンブリア爆発の規模と原因 カンブリア紀と先カンブリア時代の境の地層から、およそ数百万年の幅で、急に多細胞動物の化石が顕在化する。スティーヴン・ジェイ・グールドに代表されるように、急に化石が見つかることから、少数の祖先種から爆発的に種の多様化が起きたのだと考えられている(図1a)。一方、最近の分子からの知見を踏まえて、リチャード・フォーティのように、もっと緩やかな“爆発”を考えることも可能である(図1b)。 図1.カンブリア爆発についての2つの対立する考え a)グールドの芝生モデル.およそ5億4千万年前に、1つの祖先から一斉にいろいろな動物門が出現したとする説。 b)フォーティの説.動物の多様化はむしろ緩やかに起こり、およそ5億4千万年前頃から化石として顕在化したが、それ以前は小さく、化石として見つけにくいとする説。Xは絶滅して系統を表す。 最近の分子系統学的解析によると、多細胞動物の主要なグループである三胚葉動物は、まず脊椎動物を含む僅かなグループからなる新口動物と、多くの三胚葉動物を含む旧口動物に分かれ、新口動物、旧口動物それぞれがさらに分岐を繰り返して、現在の多様な動物門が形成された。ラッセル・ドリトルらの計算によると、新口動物と旧口動物の分岐はおよそ6億8千年前で、それは化石が顕在化した5億4千年前のカンブリア爆発よりも遙かに古い。三胚葉動物は6億8千年前より多様化を繰り返したが、この頃の動物は体が小さく、化石として残りにくく、また発見されにくい。そして最後の数百万年前より急速に大型化して、化石として残るようになったというのがフォーティの考えである。すなわち、三胚葉動物の多様化は先カンブリア時代にすでに進んでいて、最後の数百万年で大型化したため、化石として顕在化したというのだ。 フォーティの説では、多様化はグールドの説に比べて緩やかに起きていたことにはなるが、それでも6億年前の年代を中心に前後数千万年の幅で多様な動物が急速に進化したことになる。ここではフォーティに従って、動物の多様化が6億年前付近で急速に進んだと考えておこう。 ではなぜ、動物は徐々に多様化せずに、ある特定の時期に爆発的に多様化したのか。この問に対しても明快な答えはいまだにない。はっきりしていることは、カンブリア紀と先カンブリア時代の境で多細胞動物の化石が急に出現するということだけなのだ。昔からいわれている、カンブリア爆発に対する伝統的な説明は、当時の地球環境の変化に基づいている。すなわち、(1)この頃になって、多細胞動物が活発な代謝を行うために必要な自由酸素量が現在とほぼ同じ20%のレベルに達した。この自由酸素量の増加が大型動物の出現を可能にした。また、(2)オゾン層が出現して、気候が温暖化したことも要因の一つと考えられている。さらに、(3)地球の大陸と海の分布は当時と現在ではずいぶん違っていて、先カンブリア時代の終わりには、大陸は一つの大きな塊となって、超大陸を形成していたと考えられている。それが徐々に分裂を始め、それに伴って大陸棚や海の浅瀬が出現した。こうした環境は動物たちに新しい生態的環境を提供する結果となった。これは先住者も競争相手も全くいない、カンブリア紀の動物たちにとって生態的に空っぽの環境となった。競争相手のいない環境では、体のデザインや動きの俊敏さはまったく問題にならないので、奇妙な形をした動物たちでカンブリア紀の海はまたたく間に埋め尽くされていったというのだ。 最近、スノーボール仮説という、当時の地球環境に対する新しい仮説が提唱され、カンブリア爆発との関係で話題となっている。この仮説によると、当時の地球では、全地球の凍結と氷解が繰り返されていたらしい。最終氷解期では、氷の解けた浅瀬の海はカンブリア紀の動物たちに新しい生態的環境を提供する結果となったことであろう。カンブリア爆発はちょうどこの時期に起きたというのだ。 ▲このページのトップへもどる 遺伝子重複が起きた時期を推定する 上で述べたように、今から6億年ほど前に小さな体の多細胞動物が急速に多様化し、それから5千万年ほど後に、そうした中から化石として認識できるほどの体の大きなグループが爆発的に増加した。この形態レベルで爆発的多様化が進行しているまさにその時、遺伝子レベルでは何が起きていたのか?表現形レベルに呼応して、遺伝子レベルでも盛んに遺伝子重複が起き、遺伝子の多様化が急速に進んだのか? ここで考える遺伝子は、解糖系の酵素をコードしている遺伝子や、キネシン遺伝子のような全ての真核生物が共有する遺伝子ではない。そうではなく、細胞間情報伝達や形態形成に関与する遺伝子など、多細胞の動物に特有と考えられている遺伝子である。こうした多細胞動物特有の遺伝子は進化の過程で遺伝子重複を繰り返し、多数のメンバーから成る大きな遺伝子族を形成している。われわれが答えるべき問は次のようになる。それぞれの遺伝子族がカンブリア爆発に呼応して、高い頻度で遺伝子重複を繰り返し、爆発的にメンバーの数を増やして、多様化したのであろうか? 上で述べたように、この問題は、遺伝子重複が起きた時期が何時かという問題に焼き直った。時間を推定する場合、分子時計の性質を利用すると便利である。分子には、同じ遺伝子を異なる種間で比較すると、時間の経過に伴って、一定の割合で塩基ないしはアミノ酸の置換を起こす性質があることが知られている。この性質を分子時計という(本シリーズ、「パラダイムシフト:分子進化の中立説」を参照)。しかし、現在では分子時計は常に存在するわけではなく、限られた条件で成立すると考えられている。特にここでは、同じ遺伝子ではなく、遺伝子重複で作られた異なる遺伝子の分岐時期を推定する問題であるから、分子時計が使えない。やや面倒だが、別な方法に頼らざるを得ない。すなわち、生物の系統と分子の系統を同時に推定することで、遺伝子重複が起きた時期を大雑把に見積もろうという方法である。 同じ遺伝子を異なる生物種の間で比較することで、生物が進化の過程でどのように枝分かれしてきたかを示す、分子系統樹を推定することができる。生物の系統樹を推定する場合と全く同じ考えで、一つの遺伝子族に属する遺伝子の系統樹を推定することができる。分子系統樹における枝分かれは、生物の系統樹では種分岐に対応し、遺伝子族の系統樹では遺伝子重複を示す。これら2つの系統樹を同時に推定することで、遺伝子重複が起きた時期が推定できる。この系統樹をここでは複合系統樹と呼んでおこう。以下で複合系統樹から遺伝子重複が起きた時期をどう推定するかを簡単な例で解説しよう。 ヘモグロビン(Hb)は2つの異なるポリペプチド鎖、α鎖とβ鎖、それぞれが2つずつ重合した4量体(α2β2)で構成されている。α鎖とβ鎖は脊椎動物の進化の過程で遺伝子重複で作られた。いま、ヒトのα鎖、コイのα鎖、ヒトのβ鎖、及びヤツメウナギのHbの比較から複合系統樹を作ったところ、図2のようになったとしよう。ここで分岐点aは、同じα鎖の遺伝子の系統で、生物種ヒトとコイが分岐した時期に対応する。古生物学的にこの時期はおよそ4億年前と推定されている。分岐点bはヤツメウナギ(無顎類)と有顎類の分岐に対応し、およそ5億年前に遡る。分岐点cは遺伝子重複によってα鎖とβ鎖が分かれた時期に対応し、それは〜4億年前と〜5億年前の間に推定されることになる。 図2.ヘモグロビンα鎖ーとβ鎖ー遺伝子の重複時期の推定法 分岐点aは四足動物と魚類の分岐(〜4億年前)に対応し、分岐点bは有顎類と無顎類の分岐(〜5億年前)に対応する。 分岐点cは遺伝子重複による遺伝子aと遺伝子bの分岐に対応し、従ってその時期はおよそ4億年前から5億辺前と推定される。 ▲このページのトップへもどる 遺伝子族:サブファミリーと組織特異的遺伝子 多細胞動物に特有の遺伝子族として、チロシンキナーゼ(PTK)遺伝子族を例に考えてみよう。タンパク質リン酸化酵素(プロテインキナーゼ)は、互いに相同な触媒部位(キナーゼドメイン)を共通に持つ巨大な遺伝子族を形成している。ここでドメインとは、アミノ酸配列が立体構造上、かたまりをなしている領域で、通常、アミノ酸が100あるいはそれ以上の大きさからなる。一つのドメインは特有の機能をもつ場合が多い。 プロテインキナーゼは基質の違いにより、セリン・スレオニンキナーゼ(PSK)族とPTK族の2つに大きく分けられる。プロテインキナーゼ族の分子系統樹から、PTK族はPSK族の1グループから進化したことが分かっている。ほとんど全ての真核生物に存在するPSKに対し、PTK族は多細胞動物のほかは限られた系統にしか見つかっていない。細胞性粘菌にはチロシン特異的なリン酸化活性を持つものがあるが、これらは別のPSKの一グループから進化してきたもので、多細胞動物のものとは由来が違う。PTKは主に細胞間のシグナル伝達に関わり、細胞の増殖や分化を制御している。これらのことからPTKは菌類・植物から分岐後、動物の系統で進化したと考えられている。 PTKはキナーゼドメイン以外にも幾つかのドメインから構成されていて、異なるドメイン構成を持つPTKの間では基本的な機能が違っている。遺伝子重複による異なるドメインの生成とドメイン混成によって、多様なPTKが進化の過程で作られてきた。図3の右半分に各PTKメンバーのドメイン構成が示されている。 図3.チロシンキナーゼ族の分子系統樹 チロシンキナーゼ(PTK)族の分子系統樹の一部が示されている。青で塗りつぶされている系統の塊がサブファミリーに対応する。赤丸はおよそ9億年前に起きたとされる、カイメンとそれ以外の動物との分岐に対応する。赤の菱形は異なるサブファミリーを作った遺伝子重複を示す。いずれの遺伝子重複もカイメンとそれ以外の動物との分岐前に起きている。各サブファミリーのドメイン構成を図の右側に示した。 異なるドメイン構成を持った多様なPTKが進化したずっと後の時代に、各々のPTKがさらに遺伝子重複によってコピー数を増やす。ここでの遺伝子重複は遺伝子領域全体のコピーなので、コピーで増えたメンバーは互いにドメイン構成が同じである。そのため機能がほとんど同じと考えられる。このプロセスが進むと、共通のドメイン構成を持った複数のメンバーからなる遺伝子グループが形成される。このグループをサブファミリーと呼んでいる。PTK族は多数のサブファミリーによって構成され、かつ各々のサブファミリーはさらに複数のメンバーで構成されという二重構造になっている。一般に、同一サブファミリーに属する異なるメンバーは、機能がほぼ同じだが、発現する組織・器官が異なる場合が多い(こうしたメンバーのことを組織特異的遺伝子と呼ぶことにする)。PTK遺伝子族に見られるこの階層構造は、多くの多細胞動物に特有の遺伝子族に共通に見られる特徴になっている。 ▲このページのトップへもどる 遺伝子族の多様化とカンブリア爆発との関連 さおよそ10億年前、植物、菌類から動物(後生動物)の系統が分岐し、最初にカイメン(側生動物)とそれ以外の動物、すなわち、真正後生動物とが分岐する。その時期はおよそ9億年前と推定される。次いで、真正後生動物の系統で、クラゲ、イソギンチャク、ヒドラなどの二胚葉動物と主要な動物のグループである三胚葉動物とが分岐する。 三胚葉動物はおよそ6億8千年前に、まず新口動物と旧口動物のグループに分かれる。旧口動物のグループは、節足動物や線形動物を含む脱皮動物のグループと軟体動物や扁形動物を含む冠輪動物のグループに大きく分かれ、それぞれのグループはさらに次々と現存の動物門へと分かれていく。 新口動物の方は、ウニ、ホヤ、ナメクジウオを経て、脊椎動物が出現する。脊椎動物の系統では、5億年ほど前に、メクラウナギやヤツメウナギの無顎類と顎のある有顎類に分かれ、後者の系統では、4億年ほど前に魚から四足動物が進化する(本シリーズ、変わる動物の系統樹-三胚葉動物の体腔と系統分類-を参照)。 これらの動物をできるだけ含めながら、それぞれの動物から一つの遺伝子族、例えばPTK遺伝子族のメンバーをPCR法で網羅的に単離し、分子系統樹を推定する。その系統樹に基づいて、各々のメンバーを作った遺伝子重複がいつ頃起きたかを図2の方法で推定する。このプロセスを別の遺伝子族で繰り返し、多数の遺伝子族に対するデータを集積する。当時著者が属していた京都大学の研究室で、10年近くかけて、15ほどの多細胞動物特有の遺伝子族の解析を行った。そのうちの一つ、PTK遺伝子について結果を紹介しよう。他の遺伝子族についても基本的には同じ結果になる。 当時大学院生だった菅裕博士を中心に、著者の研究グループで推定されたPTK族の分子系統樹の一部が図3に示されている。この系統樹は異なるPTKの間で共通に存在するチロシンキナーゼドメインの配列比較から推定された。この系統樹の大きな特徴は、異なるサブファミリーが分岐した時期が非常に古いことである。図3の系統樹によると、異なるサブファミリーを作った遺伝子重複が、現存する多細胞動物のなかで、最も古い時期に出現したと思われるカイメン〔側生動物〕とそれ以外の多細胞動物(真正後生動物)が分岐する以前に起きている。つまり、PTKの基本的な機能の多様化はほぼ9億年前に完了していたことになる。カンブリア爆発による動物の多様化が6億年前頃に起きているので、形態の多様化と遺伝子の多様化には時間的なずれがあることになる。 ところで、図3の複合系統樹は全てのPTKが共通に持つキナーゼドメインで推定した。従って、側生動物と真正後生動物の分岐前に起きたのは多様なキナーゼドメインを作った遺伝子重複であって、ドメイン混成が起きた時期はもっと後の時代、すなわち、カンブリア爆発当時である可能性が残る。このことを検証するには、カイメンとその他の動物の間で、それぞれのPTKのドメイン構成を比較する必要がある。調べた限りドメイン構成は、同じサブファミリー内で比べると、カイメンとその他の動物で共通していた。このことは、ドメイン混成も側生動物と真正後生動物の分岐前に起きたことを意味している。 PTKの基本的な機能の多様化は動物進化の初期に完了していたという結論は他の遺伝子族にもあてはまるだろうか?PTK族以外のシグナル伝達系に関与する遺伝子族や形態形成遺伝子族など、多くの遺伝子族について、同様の解析を行ってみると、どれもPTKの多様化パターンに酷似していることが分かる。 こうして、異なるサブファミリーを作った遺伝子重複のほとんどは、ドメイン混成も含め、動物進化のごく初期、すなわちカイメンとその他の動物が分かれる以前に起きていたことになる。菌類にはこうした遺伝子のほとんどは存在しないことから、動物特有の遺伝子は、菌類との分岐の後、側生動物/真正後生動物との分岐前にほとんどの機能的多様化を済ませていたことになる。もっとも菌類で大々的な遺伝子の欠失が起きていたとすると、遺伝子の多様化はもっと古い時代にさかのぼることになる。 異なるサブファミリーを作る遺伝子重複の他に、同じサブファミリーに属するメンバー(すなわち、組織特異的に発現する遺伝子)もまた遺伝子重複で多様化した。では、このタイプの遺伝子重複はいつ頃起きたのか?PTK遺伝子族に属する一つのサブファミリーである血小板由来成長因子レセプター(PDGFR)サブファミリーでみてみよう。 図4.血小板由来成長因子レセプター(PDGFR)サブファミリーの系統樹(一部) 図4はPDGFRサブファミリーの詳しい系統樹の一部で、組織特異的遺伝子の重複は、脊索動物のナメクジウオから分岐後、脊椎動物の系統で、およそ5億年前に起きた無顎類(メクラウナギとヤツメウナギ)と有顎類が分岐した辺りで盛んに起きていることが分かる。他のいくつかのサブファミリーでみても同様の結果が得られる。 この系統解析からもう一つの興味ある情報が得られる。詳しく述べないが、これらの遺伝子が乗っている染色体上の位置から、2回の遺伝子重複の後、2回の染色体レベルの重複が、いずれも無顎類と有顎類が分岐した辺りで起きたと推定される。おそらく、一つの遺伝子族の各々のサブファミリーは、無顎類と有顎類が分岐した辺りで遺伝子重複と染色体の重複とでメンバーの数を増やし、遺伝子多様性を高めたと推定される。他のサブファミリーも同様の多様化パターンを示す。 図5.カンブリア爆発と遺伝子多様化の時期 四足動物へ至る系統を中心にした系統樹.左へいくに従って時間が遡る。 以上のことから、菌類、植物から分岐後、四足動物へ至る動物の系統でみると、遺伝子の多様化パターンは次のようにまとめることができる(図5)。およそ10億年前、菌類・植物から分岐した多細胞動物は、およそ9億年前に起きた最初の現生動物の大きな分岐、すなわちカイメンとその他の動物の分岐までのおよそ1億年の間に機能の違う、さまざまな基本的遺伝子(遺伝子族のサブファミリーに対応する)が、遺伝子重複とドメイン混成の機構で、急速に多様化した。その後、無顎類と有顎類が分岐した、およそ5億年前の時期までの4億年の間は新しい遺伝子の創造はほとんど起きていない。組織ごとに特異的に発現する遺伝子(遺伝子族の同一サブファミリー内メンバー)が、一部遺伝子重複で、一部染色体レベルの重複で無顎類と有顎類が分岐したあたりで盛んに多様化し、遅くも四足動物へ至る系統が魚類から分かれた、およそ4億年前までに多様化が完了していた。 上で述べた遺伝子多様化パターンには二つの大きな特徴がある。第一に、遺伝子は徐々に多様化するのではなく、断続的に短い期間に集中して多様化する。動物特有の遺伝子では、2つの限られた期間(それぞれおよそ一億年の間)に集中している。興味あることに、どの遺伝子族で見ても、似たような時期に多様化している。これにはゲノムレベルの大規模な重複と、繰り返し配列が特定の時期に挿入され、増殖したことによって、遺伝子重複が容易になったことと関連するのかも知れない。 第二に、これは重要な結果だが、およそ6億年前に爆発的に動物の形態が多様化したと思われる時期、すなわちカンブリア爆発、には遺伝子多様化はほとんど見られない。遺伝子レベルの多様化と形態レベルの多様化の時期は明らかに重ならない(図5)。 この遺伝子多様化パターンが示唆する最も重要な点は、遺伝子の多様化はカンブリア爆発の直接の引き金ではなかったということである。カンブリア爆発と遺伝子爆発の時間的ずれは、カンブリア爆発の分子機構を考える上で、新しい遺伝子を作るというハードの視点ではなく、すでにある遺伝子をいかに利用してカンブリア爆発を達成したかというソフトの視点が重要であることを物語る。 ▲このページのトップへもどる 立襟鞭毛虫:単細胞原生生物に多細胞用の遺伝子セット 多細胞動物に最も近縁な単細胞原生生物は立襟鞭毛虫の仲間である。立襟鞭毛虫はカイメンの襟細胞によく似ていることから、古くから両者の近縁関係が指摘されていたが、最近になって分子系統学的に裏付けられた。多細胞動物の進化の歴史に於いて最古の分岐であるカイメンとそれ以外の動物の分岐以前に多細胞特有の遺伝子の多様化が完了していたという結果は、否応なく立襟鞭毛虫を遺伝子多様性の舞台に引きずり出す。立襟鞭毛虫は単細胞生物だから多細胞特有の遺伝子族の各々について、多細胞動物と共通の遺伝子が一つあるか、あるいは全くないと期待される。前者の場合は、その一つの遺伝子が起源となって立襟鞭毛虫との分岐後、多細胞動物の進化の過程で繰り返し遺伝子重複を起こし、一つの遺伝子族へと発達したと考えられる。後者では、遺伝子重複の種になった遺伝子が立襟鞭毛虫との分岐後、多細胞動物の進化の過程で作られ、遺伝子族へと発展したと期待される。 この疑問に答えるべく筆者の研究グループでは、チロシンキナーゼ(PTK)やチロシンポスファターゼ(PTP)など、多細胞動物特有の遺伝子のいくつかを取り上げ、単細胞の立襟鞭毛虫に多細胞の遺伝子を探査した。 PTK遺伝子族の解析からスタートしたが、驚いたことに、多細胞動物特有だと思われていたPTK遺伝子が立襟鞭毛虫に多数発見された。さらに遺伝子の系統解析の結果、多様なキナーゼドメインを作った遺伝子重複のほとんどは、動物と立襟鞭毛虫との分岐以前にすでに完了していたというまったく予想外の結果となった。ドメインの混成はその分岐の前後で盛んに起きていたようで、動物と共通のドメイン構成を持つ遺伝子もあれば、動物、立襟鞭毛虫それぞれで独特のドメイン構成を持つ遺伝子もある。 われわれは、多細胞生物には多細胞独特の遺伝子があって、そうした遺伝子が多細胞らしさを形作っていると信じてきたが、その考えはどうやら捨てなければならないようだ。非常に多数の遺伝子重複が立襟鞭毛虫と動物の分岐以前に起きていたらしいので、われわれ動物が持つ細胞間情報伝達遺伝子や形態形成遺伝子は単細胞の立襟鞭毛虫にかなり多数存在していると思わねばならない。ひょっとして、立襟鞭毛虫は多細胞生物が単細胞生物に退化したのではないかと思われるほど、遺伝的には多細胞的だ。 立襟鞭毛虫ゲノムの全DNA塩基配列の情報は多細胞性の起源と進化について興味深い。立襟鞭毛虫と動物のゲノム比較から、多細胞化がどのようにして起きたのか、その分子的基礎を解明するための多くの情報が得られるであろう。現在、京都大学の岩部直之博士が中心になって、立襟鞭毛虫の全ゲノム塩基配列決定の計画が進行中である。また、立襟鞭毛虫が持っているPTKはこの生物でいったい何をしているのであろうか?大阪大学微生物病研究所の岡田雅人教授のグループは、この興味ある問題の解明に向けて研究を進めている。 ▲このページのトップへもどる 遺伝システムの柔軟性とソフトモデル ところで、冒頭で述べた、ヒトの全遺伝子数は約37000で高々ショウジョウバエの2倍ほどでしかないという報道関係者の驚きだが、確かに脳をはじめとして、両者の形態的複雑さの違いからみて、2倍という数字はいかにも少な過ぎるように思われる。しかし、生物多様性の問題は遺伝子を作るというハードの視点ではなく、既存の遺伝子をどう利用するかというソフトの視点で理解すべきであるという立場に立つと、2倍という数字にはそれほど違和感がない。 Pax6遺伝子は眼の形態形成遺伝子として有名であるが、哺乳類では、発生の初期には神経管の形成に関与する。さらに発生が進むと、眼だけでなく、顔面の形成にも関与するらしい。さらに成体では、インスリンのα細胞の誘導にも関与しているという最近の報告もある。このように一つの遺伝子が発生の過程でさまざまな用途に利用されるならば、数少ない遺伝子数で十分形態的複雑さが達成できるのかも知れない。遺伝子や遺伝子システムは機能・構造的にかなり柔軟で、一つの機能には一つの遺伝子という融通の利かないものではなさそうだ。 融通の利く遺伝子機能はまた遺伝子の欠失を容易にするであろう。生物は環境に適応し、特殊化していく過程で、多くの遺伝子を失う。生物がもとの環境に戻ったとき、失った遺伝子は戻らないが、よく似た遺伝子で急場をしのぎ、後に失った一部の遺伝子を遺伝子重複によって復活させることができるであろう。この場合、復活といっても失ったものとは配列の上でずいぶん違っているはずだ。 動物進化の初期にすでに獲得していた遺伝的多様性は、その後の環境への適応と特殊化によって、多くの遺伝子が削り取られ、ゲノムのシェイプアップが進んだ。いっぽうで、脊椎動物へ至る系統では、ゲノムの重複などでゲノムサイズが回復している。 カイメンのゲノムサイズはヒトのおよそ1/4だが、脊椎動物のゲノムはその進化の初期に4倍化したので、両者のゲノムサイズの違いは十分理解できる。しかし、ショウジョウバエのゲノムサイズはヒトの1/20で、カイメンの1/5と比べて極端に小さい。線虫もヒトの1/30なので、脱皮動物のごく初期の段階でかなりの遺伝子の欠失があったと推測される。それでも何とかやり繰りしているのは遺伝システムの柔軟性によるのであろう。 動物の種類によっては、進化の過程で環境にますます適応し、特殊化していくものもある。他の生物への寄生はその典型で、安定した細胞内環境に適応した寄生性動物は多くの組織を失い、それに伴って遺伝子の欠失を起こす。それはゲノムサイズの縮小をもたらし、結果として子孫を増やすことに寄与する。遺伝システムの柔軟性は必要以上にゲノムを短縮することを可能にするであろう。それはますます子孫の増殖をもたらすに違いない。 魚や環形動物に寄生するミクソゾアは、単細胞にまで退化が進んだ多細胞動物である。これまで単細胞原生生物として分類されていたが、最近の分子系統学的解析によって多細胞性が明らかになった。この動物の全DNA塩基配列の情報は興味深い。いかなる遺伝子が削り取られ、いかなる遺伝子が代用しているのか、明らかになるであろう。 生物の擬態ほど生命現象の不思議さ感じさせるものはないであろう。その擬態こそ遺伝システムの柔軟性に基づく多様化のソフトモデルがぴったりな現象なのかも知れない。擬態されている生物(モデル)と見た目にはほとんど区別がつかない形態が、全く予想外の遺伝的材料を利用することで達成されているように思われる。もしそうなら、それは多様性のソフトモデルのまたとない証言者となろう。 ▲このページのトップへもどる 形態進化の便宜主義と遺伝子進化のソフトモデル 互いに外見がよく似た生物が、実は系統的にかけ離れたグループに属することを知ったとき、生物の不思議さをつくづく感じる。由来の異なる生物間に見られる外見上の類似性は、似たような生活様式に適応した結果である。クジラが水中を泳ぐという生活に適応した結果、魚によく似た流線型の形態に進化したという説明は誰でも知っている。この類似性のことを収斂と呼んでいる。有袋類と有胎盤類とでは多くのグループの間で形態上の類似性が認められる。 スティーブン・ジェイ・グールドは彼の著書、「パンダの親指」のなかで、有袋類のフクロオオカミと有胎盤類のオオカミが示す収斂現象に基づいたアーサー・ケストラーの反ダーウィニズムの考えを紹介している。ランダムな突然変異と自然選択だけで一つの生物の進化を考えることが困難なのに、ましてやよく似た生物が独立に進化する収斂現象は、いわば奇跡を二乗するようなものだとケストラーは主張する。グールドの反論を待つまでもなく、収斂は決して奇跡的な出来事ではない。2つの生物は細部に至るまでよく似ているわけではなく、類似はむしろ表面的にすぎない。だからこそ分類学者は共通の由来に基づく分岐と収斂とを容易に区別できるわけである。もっとも最近分子系統樹から明らかにされた大陸間にみられる大規模な有胎盤類の収斂については、長い間正しく理解されることはなかったのだが(本シリーズ「収斂進化2:真獣類の大陸内適応放散と収斂進化」を参照)。表面的にせよ、きわめてみごとな形態的類似が進化のタイムスケールでみて短期間に進化することが収斂の大きな不思議であり、それを可能にするメカニズムの解明は、誰にとっても興味ある、重要課題であろう。 収斂は、おそらく擬態も同じと思われるが、表現形進化の大きな特徴である便宜主義のあらわれである。生物は、ある問題を解決するために、形態や器官を本質に戻って一から組み立てることをしない。まわりにある材料を手当たり次第に利用する。素材は何でもかまわない。問題の解決のしかたはまったく便宜的である。例えば、飛揚という問題を解決するために、鳥とコウモリはそれぞれ独立に前肢を翼に進化させた。けっして一から翼を作ることをしていない。天使は背中に翼を持っているが、これこそ神?の創造であって、生物進化の所産ではない。 上で述べたように、多細胞動物の爆発的多様化においては、わざわざ新たな多細胞動物特有の遺伝子が作られたわけではなく、既存の遺伝子を利用して達成したというのがソフトモデルであった。これは分子レベルでの便宜主義とみることが出来よう。分子レベルの便宜主義が表現形進化の便宜主義とどう関わっているのか?すなわち、ソフトモデルは表現形進化の分子的基礎を与えることができるであろうか?収斂や擬態の研究からソフトモデルにそった遺伝子の利用法が具体的に明らかにされるものと期待したい。 ▲このページのトップへもどる [宮田 隆] 宮田 隆の進化の話 最新号へ