年度別活動報告
年度別活動報告書:2002年度
イチジク属とイチジクコバチの共進化
蘇 智慧(研究員、代表者)
東 浩司(奨励研究員)
神田嗣子(研究補助員)
中村桂子(館長)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物は陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(競争、共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの共生関係についてDNA解析や野外調査により研究を行っている。
イチジク属Ficusは、クワ科(Moraceae)に属し、4亜属(イチジク亜属Ficus, アコウ亜属Urostiguma, Pharmacosycea, Sycomorus)に分けられ、熱帯を中心に約750種が世界中に分布している。日本には南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。一方、イチジクコバチの仲間は分類学的に膜翅目、コバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとして分類されている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つは送粉コバチ亜科Agaoninae (pollinating fig-wasps)で、他の亜科はすべて寄生コバチによって構成されている1)。送粉コバチとイチジクとの間、子孫を残すために互いに必要としており、「1種対1種」という厳密な対応関係を持っていると言われている。しかし、寄生コバチの場合は、複数種の寄生コバチが同種のイチジクに寄生することができる。イチジクとイチジクコバチとの関係は古くから共進化のモデル系として多くの研究をなされてきた。特に近年アフリカや南米の材料を中心に分子系統解析の研究も盛んに行われるようになった2)。日本産イチジク属とイチジクコバチについても研究が行われている3, 4)。
我々は昨年度メキシコ産のイチジクとイチジクコバチの分子系統解析を行ったところ、同じ種のイチジクから採集した送粉コバチが系統樹上1つの枝にまとまらず、異なる系統に分かれるという結果が得られた。異なる起源のコバチが同種類のイチジクに送扮しているらしい。この現象は日本産イチジク属にも起きているのか、「1種対1種」の関係はどれ程厳密に維持されているのかを調べるために、今年度は日本産イチジクを材料にして各地からイチジクの葉とそのコバチを採集し、葉緑体DNA、核rRNA遺伝子、ミトコンドリアCOI遺伝子を用いて分子系統解析を行った5)。また日本産イチジクとイチジクコバチの地域的変異も調べた。
結果と考察
1) 日本産イチジク属と送粉コバチの分子系統解析
イチジク属の系統解析の結果、日本産イチジク属は5つのグループに分けられることが示された(図1左側A-E)。これらのグループのまとまりは分類学・形態学的にも支持されるものである.簡単に説明するとムクイヌビワF. irisana・ホソバムクイヌビワF. ampelas・ハマイヌビワF. virgata(図1-A)は小高木または灌木で小さい堅い花嚢を付ける。オオイタビF. pumila・ヒメイタビF. thunbergii・イタビカズラF. nipponica(図1-B)はつる性のイチジク属である.オオバアコウF. caulocarpa・アコウ・ガジュマル(図1-C)は雌雄同株であり(他の日本産イチジク属はすべて雌雄異株)、大木になり、直径8mm程度の柔らかい花嚢を葉腋や幹に直接つける。特にガジュマルは気根を垂らし絞め殺し植物として知られている。オオバイヌビワF. septica・アカメイヌビワF. benguetensis・ギランイヌビワF. variegata(図1-D)は比較的大きい葉を付け、堅いごつごつした直径2cm内外の比較的大きい花嚢を付ける。イヌビワF. erecta(図1-E)は単独で一つのグループを形成し、日本産のどの仲間と近いのか現時点では不明である。イヌビワの個体はあまり大きくならないが、花嚢は熟すと直径2cm程度の大きさになる。ムク・ホソバムク・ハマイヌビワのクレード(図1-A)を除いてこれらのグループの単系統性は高いブートストラップ値(95%以上)で支持された。ムクイヌビワとホソバムクイヌビワ、アコウとオオバアコウはそれぞれ葉緑体DNA塩基配列に違いが見られず、遺伝的にほとんど分化していないことが示唆された。種内で塩基配列に違いが見られたのはアカメイヌビワ、ギランイヌビワ、イヌビワ、オオイタビで、特にイヌビワは沖縄本島以北のサンプルが一つのクレードにまとまり、沖縄本島と宮古島との間で分化が生じていることが示唆された。
イチジク属の花嚢から採集した送粉コバチの核28S rRNA遺伝子の塩基配列による分子系統樹を図1の右側に示した。中央の横棒はそれぞれ対応するイチジク属と送粉コバチのサンプルを示している。アコウコバチPlatyscapa ishiianaとガジュマルコバチEupristina verticillataを除いて(図1-C)、ある1種類のイチジクから得られた送粉コバチの塩基配列には全く違いがないことが示された。逆に言うと、ある種のイチジクの花嚢に複数の異なる系統群に属する送粉コバチが現れなかったと言える。このことはイチジク属と送粉コバチがかなり厳密な1種対1種の関係を維持していることを示唆している。また、イチジク属で見られた5つのグループと対応して送粉コバチも5つのグループに分けられた。これは少なくとも近縁種間ではイチジクと送粉コバチは共進化してきたことを意味している。しかしながら、それらグループ間の系統関係となると、特に送粉コバチの系統樹ではほとんどの枝が50%以下のブートストラップ値を示したため、現時点では比較・考察することは困難である。もし、ここで得られた系統樹の樹形が正しいとすると、両者のグループ間の系統関係は明らかに矛盾しており、興味深いところである。
日本産のイチジク属と送粉コバチの系統解析の結果を紹介したが、これは世界に約750種もあるイチジク属のほんの一部であり、これをもってイチジク属と送粉コバチの共進化を論じるのはなかなか難しい。現時点で分かったことは、少なくとも日本産のイチジク属については、かなり厳密な1種対1種の関係を保持していること、また、近縁種間ではイチジク属とイチジクコバチは共進化をしてきたことが示唆されたことである。さらに、イチジク属、送粉コバチともに南西諸島の島間での分化がほとんど見られないことが分かった。多くの種が分布している熱帯のイチジク属でも同じような結果が得られるかどうかは今後の課題である。
|
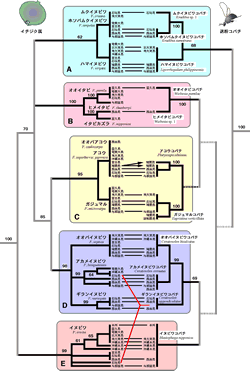
図1 イチジク属と送粉コバチの系統樹
2) 送粉コバチと寄生コバチの分子系統解析
イチジク属の花嚢からは必ずと言っていいほど、送粉コバチ以外のコバチ(寄生コバチ)が見つかる。すでに述べたが、分類学的には送粉コバチは1つの亜科で、寄生コバチは複数の亜科からなる。しかし、両者間の系統関係は明らかではない。以下3つの仮説が考えられる。(1)イチジクコバチは複数の系統(亜科?)に分かれ、そのうちの1つは送粉コバチの系統で、寄生コバチは多系統または側系統群となる。(2)送粉コバチと寄生コバチは最初から二分岐し、それぞれ別の単系統群を形成している。(3)複数の寄生コバチの系統群において、送粉コバチは独立に複数回進化した。つまり送粉コバチは多系統群である。今回の系統解析に用いた28S rRNA遺伝子の断片は、送粉コバチと寄生コバチをそれぞれ比較してみると、寄生コバチ内では788~809塩基対で長さの違いは比較的小さいが、送粉コバチ内では845~912塩基対で比較的大きな長さの違いが見られた。また、送粉コバチの塩基配列は、寄生コバチのものより長いことも分かった。塩基配列の構造から見れば送粉コバチと寄生コバチはそれぞれ別系統のようである。一方、塩基配列を用いた系統解析の結果、日本産の送粉コバチと寄生コバチに限ってのことだが、それぞれのグループは単系統群を形成し、最初に両者が分岐することが示された。この結果は上述(2)の仮説を支持することになるが、系統樹にはイチジク属の4亜属のうち、アコウとイチジク 亜属の2亜属から採集されたイチジクコバチしか含まれていないため、結論はできない。そこで、われわれは最も祖先的と言われているPharmacosycea亜属のメキシコ産14種から採集された送粉コバチと寄生コバチの28S rRNA遺伝子の塩基配列をこの系統樹に加えて解析を行った。その結果、やはり明らかに最初に送粉コバチと寄生コバチが分かれ、それぞれ単系統を形成した。但し、寄生コバチの同定結果が出るまでは結論が待たれる。
送粉コバチの場合、ある種のイチジク属から得られたコバチはすべて単系統群にまとまるのだが(図1)、寄生コバチでは複数の別種と思われるコバチが1種類のイチジクから得られた。寄生コバチの種の同定はほとんど進んでいないが、例えば、ガジュマルからは12の別種と思われる寄生コバチが得られ、それらは7つの系統群に分かれた(図2)。それ以外のイチジク属の種からも複数の異なる系統の寄生コバチが得られている。このことは送粉コバチと違って、寄生コバチでは比較的簡単に宿主の転換が行われていることを示している。
3) 寄生コバチの分子系統解析
寄生コバチには長い尾(産卵管)を持つタイプと短い尾を持つ(または尾が無い)タイプがある。これら二つのグループを系統樹上に配置してみると、送粉コバチは尾が短いタイプなので、イチジクコバチ全体としては尾が短いのが祖先的(原始的)であると考えられる(図2)。そして、その中から尾が長いタイプが1回だけ進化し、さらに、再び尾が短いタイプが出現したことが見て取れる。もしくは、尾の長い系統が2回進化した可能性も考えられる。いずれにせよ、尾が長いタイプはイチジク花嚢の外から産卵できるため、明らかに花粉の運搬には寄与しない。しかし、尾が短いタイプは産卵するために花嚢内部に侵入すると思われ、送粉に全く寄与していないのか分からない。もしかしたら、ある程度は送粉に寄与しているかもしれず、イチジク属と送粉コバチの共進化機構を解明する上で、寄生コバチの生態を知ることはなにか重要なヒントを与えてくれるかもしれない。
また、系統樹から寄生コバチは宿主の転換によって種分化が起きていることが見て取れる。例えば、イヌビワオナガコバチとガジュマルオナガコバチは明らかに祖先を共有しているが、両種の分岐は勿論イヌビワとガジュマルの分岐とは関係なく遙かに新しい。つまり両種の祖先が最初にイヌビワとガジュマルのどちらか片方に寄生していて、もう片方に寄生するようになるにつれて種分化が起きたと考えられる。
|
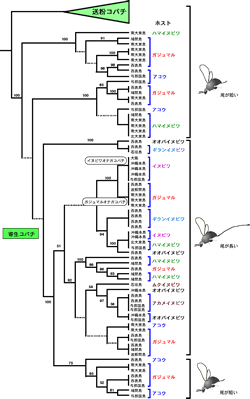
図2 寄生コバチの系統樹
おわりに
このように、イチジク属とイチジクコバチの関係は実は二者間だけの問題ではなく、寄生コバチを含めた三者の関係が絡み合った複雑な系である。イチジク属と送粉コバチの共生関係の進化を探る上ではこれら三者の生活史がどのように関連しているかを知る必要があるが、まだほとんど分かっていないのが現状である。寄生コバチは宿主の転換によって種分化を起こしているようだが、送粉コバチはイチジクとの「1種対1種」の関係がかなり安定している。送粉コバチと極めて厳密な「1種対1種」の共生関係を常に維持しながら、イチジク属はどうやって700種をこえるまでに繁栄してきたのだろう。今後 DNA 解析と生態調査などを通してこの謎を解いていきたい。
引用文献
1) Rasplus J-Y, Kerdelhue C, Clainche I L, Mondor G (1998) Molecular phylogeny of fig wasps Agaonidae are not monophyletic. C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 321: 517-527.
2) Weiblen, G. D. (1999) Phylogeny and ecology of dioecious fig pollination. Ph. D. Thesis, Harvard Univ., Cambridge, Massachusetts.
3) Yokoyama J (1995) Insect-plant coevolution and speciation. In R. Arai, M. Kato, and Y. Doi [eds.], Biodiversity and evolution, 115-130, National Science Museum Foundation, Tokyo, Japan.
4) Yokoyama J, Iwatsuki K (1998) A faunal survey of fig-wasps (Chalcidoidea: Hymenoptera) distributed in Japan and their associations with figs (Ficus: Moraceae). Entomol. Sci. 1: 37-46.
5) Su Z-H, Imura Y, Osawa S (2001) Evolutionary discontinuity of the Carabine ground beetles. J. Mol. Evol. 53: 517-529.