
脊椎動物の「かたち作りの進化」を、細胞増殖と分化そして組織移動の観点から、
カエル・イモリ・プラナリアを用いて考えています。
研究内容
1・両生類の原腸形成機構の解析
私たちは、10種以上の両生類種を用いて実験を行なうことで、両生類の原腸形成運動は前世紀初頭から信じられ、日本でも戦前から高等学校で教えられているものとはまったく異なることを示しました。
両生類の原腸形成過程の研究は、Spemannらによるランドマーク的発見に始まります。Spemannたちは、初期原腸胚の原口背唇部を、別の胚の反対側に移植することで新たな体軸の形成が誘導されることを見いだし、移植片は中軸中胚葉(脊索)にしかならず、移植された小さな組織片が本来お腹を作る予定であった細胞群に働きかけて脳など複雑な神経組織を含む背側構造を作り出すことから、この原口背唇部を「オーガナイザー」と名付けました(図1)。現在信じられているモデルによれば、正常発生においてオーガナイザーは、原口と呼ばれる裂け目を通って内部に潜り込み、それが「胞胚腔の屋根」の部分を裏打ちしながら動物極方向へとさかのぼります。オーガナイザーは、さかのぼりながら「胞胚腔の屋根」を神経組織へと分化誘導します。オーガナイザーがたどり着いたところが将来の頭部となり、したがって頭尾体軸は尾部から頭部へと形成されることとなります(図2)。
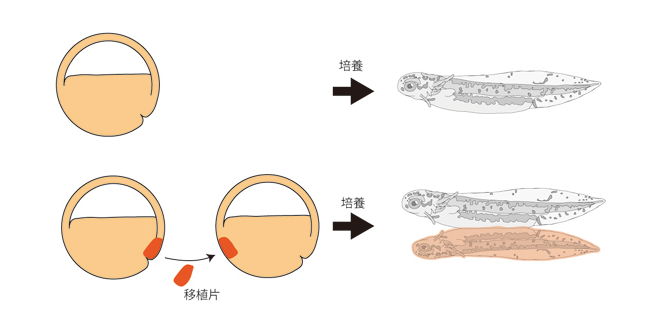
図1
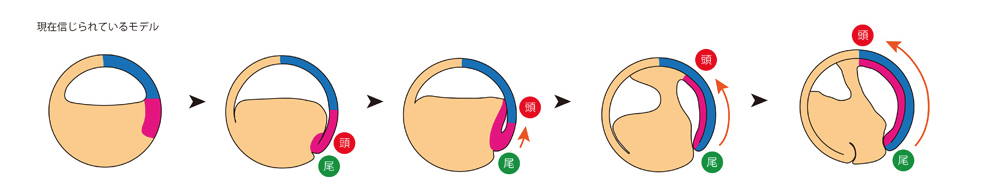
図2
しかし、私たちが提出しているモデル(図3)では、これらの相対的な位置関係や運動の方向がまったく異なります。まず、将来のオーガナイザーは胞胚期の「胞胚腔の屋根」の部分に生じます。その後、オーガナイザーは覆いかぶせ運動によって赤道方向におりてきて、初期原腸胚期期までに胞胚腔の床にかけて広がります。その後短時間のうちに胞胚腔の床にあるオーガナイザー組織は、胞胚腔の屋根が下がってくる動きに押されて、「屋根」との間に谷を作るように動き(この動きが大陸プレートの潜り込みにも似ていることから「サブダクション(潜り込み)」運動とよびます)、続いて「谷底」からゆっくりと谷を閉じるように「屋根」と「床」が接し始めます(この動きはあたかもジッパーを閉じる動きを思わせることから「ジッパリング」と名付けました)。
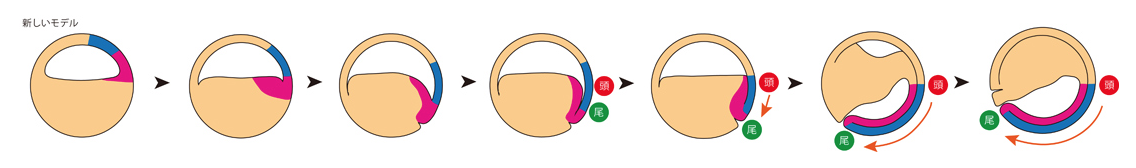
図3
このサブダクション&ジッパリング(S&Z)運動により、オーガナイザーと予定神経組織の接触が確立するということです(図4)。S&Z運動によって互いに接したオーガナイザーの最前方部と頭部神経は、その後の原腸形成過程を経ても互いにずれることなく接触し続けます。したがって、体軸は植物極を通り腹側領域へと後ろ方向に形成されることとなります。
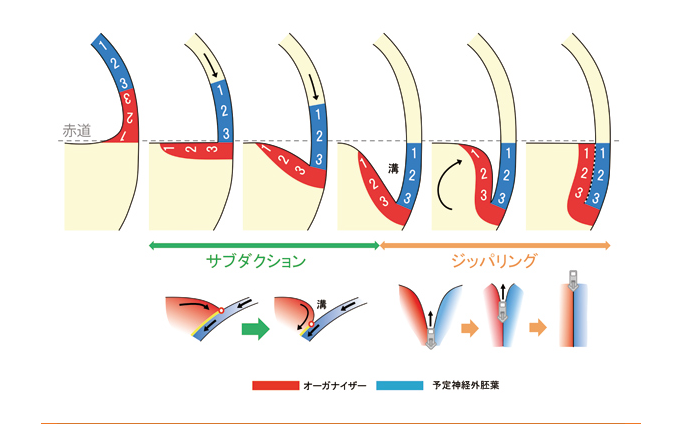 図4
図4
このモデルと従来のモデルを比較しますと、いくつかの大きな違いが見て取れます。ひとつは前方部オーガナイザーが頭部神経と接し、将来の頭部の領域が決まる場所は初期原腸胚の赤道領域であり、後期原腸胚の動物極ではないこと、もうひとつは、オーガナイザーは頭尾体軸を前方へと作るのではなく、原口背唇部が下がりながら後方へと体軸を形成することです。さらに、オーガナイザーが原口背唇部にはなく、胚の内部(胞胚腔の床)に存在していたことです。もちろん、この「胞胚腔の床」部分は、別の原腸胚の腹側に移植することで二次体軸を生じさせるというSpemannのオーガナイザーと同様の活性を有することは確認しています。
この新しいモデルを見ると、脊索動物門の原腸形成機構全体の統一モデルの確立もできそうです。まず、脊椎動物に最も近いとされる原索動物の一種ナメクジウオをみましょう。ナメクジウオの初期原腸胚は一層の細胞から成る中空のボールで、軟式テニスボールを親指で押し込むようにと表現される動きで、両生類のオーガナイザーに相当する領域(脊索)が予定神経細胞へと接して神経誘導や体軸形成を行ないます。両生類とほとんど同じ動きをしていることが分かります(図5)。
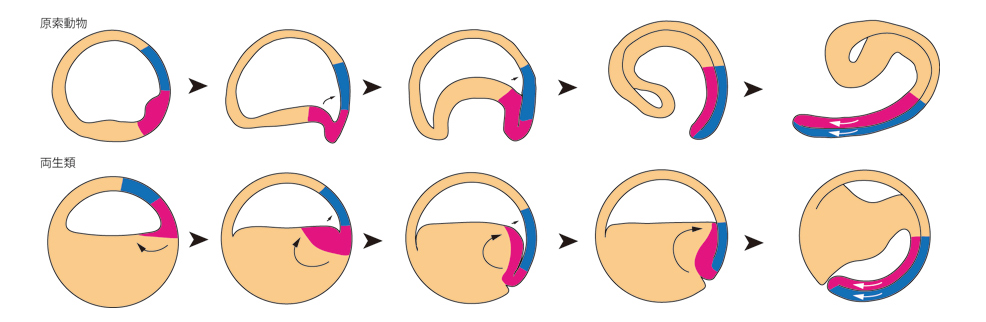
また、両者の違いは卵黄(栄養分)を持つことによって卵が巨大化した結果と考えることができそうです。さらに、これまでは全く違うとされてきた鳥類(羊膜類)ですが、両生類では神経を含む背側体軸が植物半球に形成されますので、両生類の模式図を上下逆転させるとニワトリの原腸形成とまったく同じ動きが再現できます(図6)。両生類の植物半球は卵黄を多く持つことから、ニワトリの卵黄と相同だと考えればますます両者は同じ動きに見えることとなりました。これまでの両生類の原腸形成過程を示すモデル図ではこのような比較はできず、両生類は「進化の袋小路」に入ったとすら言われてきたのですが、この新しいモデルによって、脊索動物の原腸形成を、両生類を中心として理解する可能性が高くなってきました。
私たちのモデルにより多くのことが明らかになってきましたが、逆にこれまで信じられてきたことの説明ができなくなってもいます。両生類に関していえば、たとえば、原口そのもの(ボトル細胞)は頭部を越えて前方に進みますが、その動きがどうなっているのか分かりません。また、真骨魚類(ゼブラフィッシュやメダカなど)は、このモデルからかなり逸脱していることが分かり、真骨魚類になった時点で何が起こったのかについて進化の視点から考える必要があるようにも感じます。両生類をモデルとしてこれらの問題点を整理し、脊索動物に共通する原腸形成運動を正しく記載する必要があります。
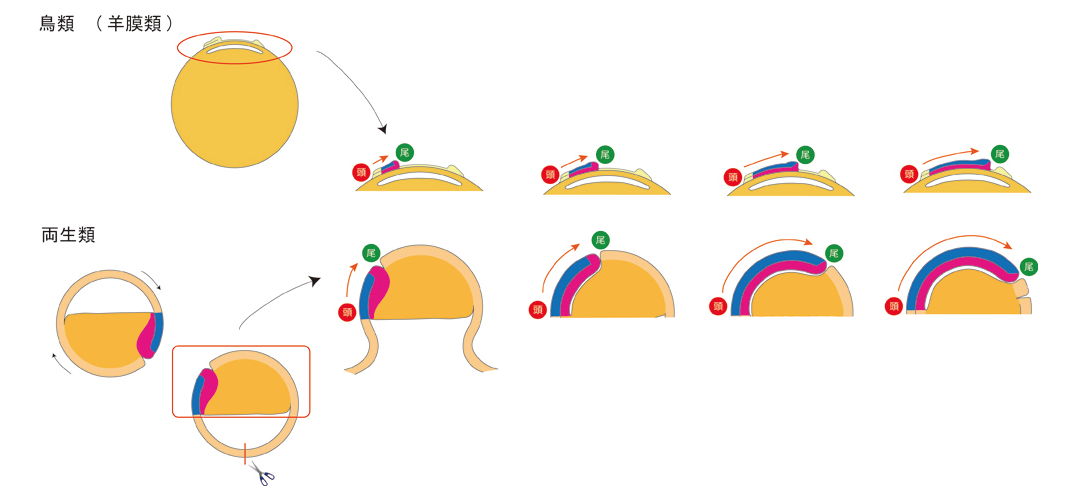
図6
2・神経堤・プラコードの形成機構を探る
脊椎動物と無脊椎(原索)動物を区別する方法の一つとして、「真の頭部」の存在が言われています。神経堤とプラコードは、頭部骨格・頭部神経節・頭部筋肉・間充織・眼鼻耳などの感覚器・末梢神経など「真の頭部」の大部分を作ることから「脊椎動物を定義する組織」とも言われ、実際に神経堤細胞とプラコードは脊椎動物以外からは見いだされていません(図3)。
神経堤とプラコードは、神経と表皮の境界領域に形成されます。これらの細胞は発生がある程度進むまでは分化をする事なく、後期神経胚期から咽頭胚期にかけてその場所に応じた分化をすることで様々なかたち作りに貢献する事から、脊椎動物の体づくりの研究において重要な位置を占めて来ました。神経堤とプラコードの形成には細胞の状態(増殖か分化か)の制御が重要であることが我々の研究から示されています。この考え方は転写制御のカスケードのみによって細胞運命が決まるという一般的な考え方に加え、そのカスケードが働ける細胞内条件(あるいはコンピテンスと言ってしまっても良いかもしれません)の重要性に言及すると同時に、神経堤・プラコードが進化的にどのように獲得されてたのかについて探る手がかりになると思います。現在は、細胞の状態を制御する機構の解析とともにその意味についても、神経堤とプラコードの領域で働いている複数の新規遺伝子機能を指標に解析を進めています。神経堤とプラコードが形成される仕組みを理解する事で、脊椎動物が進化の過程でどのように生じたのかを知る事ができると期待して研究を進めています(図4)。
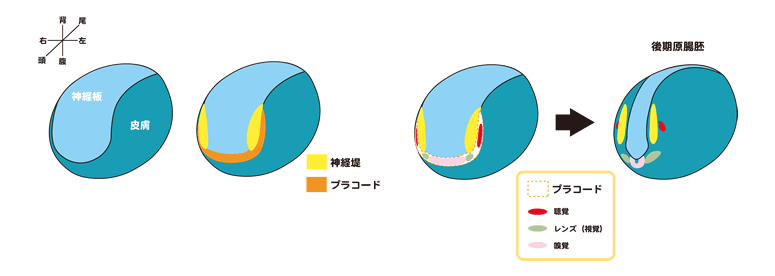

図3
メンバー一覧はこちら


最新の論文一覧はこちら
Kashima M, Komura R, Sato Y, Hashimoto C, Hirata H
A resource of single cell gene expression profiles in a planarian Dugesia japonica
Develop. Growth Differ
これまでの学位取得者とそのテーマ
- ◼︎平成28年度 修士論文 國本拓也
- 「両生類の背側構造形成に必要な最小領域の決定」
- ◼︎平成26年度 修士論文 楊 貴成
- 「両生類に普遍的な原腸形成モデルの構築〜新しい組織運動subduction & zipperingの提唱〜」
'A brand new model of amphibian gastrulation; involvement of new movements "Subduction & Zippering"' - ◼︎平成25年度 修士論文 南野怜香
- 「両生類における原腸形成運動の解析」
- ◼︎平成25年度 修士論文 西田 遥
- 「アフリカツメガエルの発生過程におけるP2Y受容体の発現パターンと機能解析」
- ◼︎平成24年度 修士論文 伊藤賢太
- 「アカハライモリの解析から見えた両生類原腸形成に共通なモデル」
- ◼︎平成23年度 修士論文 益田真都香
- 「ヌクレオチド受容体P2Y4およびP2Y11は、アフリカツメガエル初期発生過程において神経堤の維持に必要である」
- ◼︎平成20年度 修士論文 川辺実季
- 「アフリカツメガエル初期胚におけるミトコンドリアタンパク質UCP2の発現パターンと役割を探る」
- ◼︎平成20年度 博士論文 村戸康人
- 「アフリカツメガエル胚の頭部感覚器官形成におけるXhairy2の機能解析:特にレンズ形成におけるその必要性について」
"Functional analyses of Xhairy2 in the formation of cranial sensory organs in Xenopus laevis embryos: Requirements for lens formation." - ◼︎平成19年度 博士論文 永友寛一郎
- 「アフリカツメガエルの神経堤・前方神経領域形成におけるXhairy2の役割」
- ◼︎平成17年度 博士論文 山口真未
- 「アフリカツメガエル原腸形成期のオーガナイザー組織におけるXhairy2bの役割」
- ◼︎平成17年度 修士論文 村戸康人
- 「Xenopus hairy2bがオーガナイザーを領域化する二つの分子機構について」
- ◼︎平成16年度 博士論文 辻 咲織
- 「コリプレッサーGrouchoによる転写制御がアフリカツメガエルの頭部神経系の領域化に必要である」
- ◼︎平成15年度 修士論文 重永 幸
- 「アフリカツメガエル初期発生過程におけるGrouchoの機能を探る」
- ◼︎平成14年度 修士論文 山口真未
- 「鳥類胚における体壁筋形成の部域特異性」