年度別活動報告
年度別活動報告書:2002年度
脊椎動物の頭部神経系はどのように部域化されるのか
橋本主税(主任研究員)
辻咲織(大学院生)
山口真未(大学院生)
重永幸(大学院生)
皐裕美(研究補助員)
はじめに
脊椎動物の頭部構造は、頭部神経系に見られる特異的な分節構造やその形成過程など、基本的には見た目にも種を越えてよく保存されていることが分かります(1)。さらに、頭部形成を支配する遺伝子も相同の遺伝子が使われており、魚類からヒトに至まで進化的に共通の機構によって頭部の形成はなされていると考えられています(2)。しかし、基本構造は保存されているものの、例えば魚類とヒトの脳を比較すると実際の大きさや、形、あるいは各部の大きさの比率など明らかに異なっていることも事実です。これら、頭部構造の形成を制御する機構のうち、どのような共通性によって脊椎動物間での類似性が保たれ、またどのような相違点によって種間における差異が生じるのかについて明確な足掛かりを得ることが本研究のねらいなのです。この目的を達成するために両生類であるアフリカツメガエル(以下ツメガエル)の頭部形成機構を形態学と分子生物学の両面から詳細に解析しました。
両生類も含めて脊椎動物における頭部形成制御の分子機構は基本的に共通であると信じられていますが、現在までに知られている両生類の頭部形成過程は形態学的に見て他の脊椎動物種と比べてその発生様式が大きく異なっているために、「相同の分子が相異なる発生様式の中でどのように働き相同の構造を規定するのか?」という質問が生じます(3)。この質問の解答を明確にすることで脊椎動物種間での分子機構と発生機構との関係を解析するのに良い材料と考えられます。さらに両生類は、頭部形成過程の研究の歴史的な蓄積がある上に、他の種と比べ発生研究の材料としての数々の利点があるために、他種の頭部形成制御機構との相似性・相違性を明確にすることによって、種間における共通点と相違点を組織レベルと分子レベルにおいて明らかにできることが期待されます。
また、繊細かつ高度に領域化された脳として頭部神経系が形作られる機構を明らかにすることは、それぞれの脳領域が持つ独立した機能を理解し、それらの統合としての脳機能を理解する上で重要なことです。現在までに、発生の様々な過程で脳の領域化に関与すると考えられる転写因子は多数同定されていますが、それぞれ機能についての単独に解析が進んでいるに過ぎず、全体として脳が厳密に領域化される機構についてはまだ全く理解されていません(4)。この問題を解決するために、脳の領域化を制御する転写機構のネットワークの解明を目指して研究を進めてきました。
結果と考察
1)頭部領域の決定
脊椎動物の頭部領域は原腸形成期に決定されます。原腸形成過程における頭部領域の特異化には、内胚葉組織である頭部オーガナイザーと予定頭部神経外胚葉組織の物理的相互作用が重要な役割を担っています。両生類以外の脊椎動物種においては、頭部オーガナイザー組織は初期原腸胚で予定頭部神経外胚葉領域と物理的に接して存在することが知られていますが、両生類では頭部オーガナイザー組織と予定頭部神経外胚葉が物理的に接触するのは原腸形成の中期から後期にかけてであると言われています。しかし、その正確な時期は全く理解されておらず、このような発生様式の違いを起こさせる機構を明らかにすることによって脊椎動物の頭部形成を制御する普遍的な機構を求めることが重要でしょう。
そこでまず、ツメガエル胚において頭部神経と頭部オーガナイザーの物理的な相互作用が起こる正確な時期を明らかにしました。未分化外胚葉組織と頭部オーガナイザー組織が接する領域を生体染色することにより詳細に解析したところ、他の脊椎動物と同様に物理的相互作用は原腸形成期の非常に初期(ステージ10.25)にはすでに確立していることが示されました。この結果はケラーサンドイッチと呼ばれる外植体を用いた詳細な解析からも支持され、これまで予定神経外胚葉組織であると信じられてきた組織は将来の表皮であるということが明確に示されました。これらの結果から、オーガナイザー組織はこれまで言われているように尾部から頭部へと胚内部を遡りながら体軸を形作るのではなく、まず頭部が決まった後に尾部方向へと体軸を形成させるモデルを提示できました(5)。この視点に立つと、脊椎動物間での頭部体軸形成過程に組織運動のレベルで大きな共通点が見いだされます。見かけ上全く異なる運動様式によって頭部が決まると思われていた両生類が他の脊椎動物種と共通の機構によって頭部形成がなされることが明らかとなったことは、初期発生学の実験材料として最も使いやすいツメガエルの知見を元に脊椎動物一般に適応可能な統一機構の研究が進むことを意味します。
●ツメガエルの原腸形成運動を説明するモデル
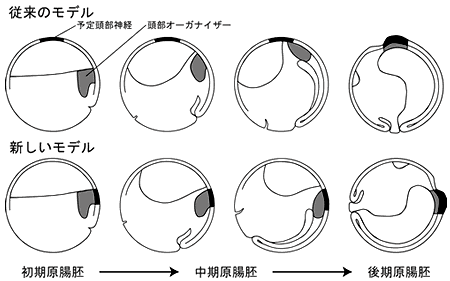
20世紀初頭以来、ツメガエルを含む両生類の予定頭部神経外胚葉は中期原腸胚まで胞胚腔の屋根にあたる部分に存在し、原腸形成期の中期から後期にかけて頭部オーガナイザー組織が予定頭部神経外胚葉領域に達すると考えられてきました。これは外部形態を間接的に標識することによって確立されたモデルでした。しかし今回、原腸胚の内部を標識する新しい方法を駆使することにより、初期原腸胚においてすでに両者は物理的に接していることが明らかとなったため、少なくともツメガエルでは他の脊椎動物と同様に、頭部領域は初期原腸胚で決まり、体軸は尾部方向に伸長することが分かりました。脊椎動物に共通する頭部形成機構の研究が始まったのです。
2)頭部神経系のパターン形成機構
脳の領域化に関わる転写因子群の多くが転写補助因子Groucho(6)と相互作用する事実に着目して解析を進めました。まず、Grouchoの機能を生体内で特異的に阻害するペプチド配列の同定を試みたところ、hairy関連遺伝子産物のカルボキシル末端に存在するWRPWモチーフが生体内のGrouchoの機能を特異的に阻害することを突き止めました。このペプチドを用いたGrouchoの機能阻害が脳の領域化に、前脳・中脳の領域化の変化、神経堤細胞の分化パターンの異常や三叉神経節形成異常など多岐にわたる莫大な影響を及ぼす事を見いだしました。この異常は、神経の部域化が具体的に進行するよりもかなり早期に、すなわち未分化外胚葉が頭部神経へと誘導を受ける時期に起こる様々な遺伝子機構の制御異常によります。ニワトリ胚では技術的な制約によって神経形成期の後半にならないと実験操作を行なえません。したがって同様の実験操作をニワトリ胚で行なったときに、ニワトリ胚の頭部神経系は見かけ上正常に発生します。
この様な総合的な異常を一分子の機能阻害が引き起こす例は国内外を問わず他に報告がなく、現在もこの研究をさらに継続しています。事実として、現在までに単離・解析がなされている脳形成に関与する転写因子において、そのほとんどにGroucho結合モチーフが存在し(7)、Grouchoによるグローバルな調節が行なわれていることは強く示唆されるにも関わらず、転写因子は全て単独の分子機構としてのみ解析が進んでいるために、同時期に同領域で起こる転写制御機構の全体像の解明には至っていません。今回見いだされたGrouchoの機能阻害による総合的な異常は、グローバルな調節を全体的に阻害したことを示している。したがってこの現象に関わる分子機構をそれぞれ素過程として抽出し解析した後、再度統合することで脳が形作られる過程に働く転写制御機構の全体像が見えてくると期待できます。
おわりに
頭部神経系の部域化に頭部オーガナイザーと呼ばれる内胚葉組織の役割が大きい事が知られています(8)。ツメガエルにおいて、頭部形成に重要な働きをする「頭部オーガナイザー」と、体軸形成に重要である「体幹部オーガナイザー」は、初期原腸胚において非常に近い領域に形成された後、それぞれ将来の頭部と体幹部の領域へと移動します。それぞれの領域を誘導する機構はかなり解析が進んでいますが、隣り合う細胞が頭部オーガナイザーとなり体幹部オーガナイザーとなる区別化の機構や、いったん誘導されたそれぞれの組織が他方の組織を誘導するシグナルによって分化転換を起こさない「性質の維持」の機構については全く解析が進んでいません。頭部神経系形成は頭部が誘導されるもっとも初期に行なわれることが今回示されましたので、頭部オーガナイザーが体幹部オーガナイザーと明確に区別される機構の解明は頭部神経形成を解明する為にも重要です。
Grouchoが脳の領域化機構の全体に影響を及ぼしていることは本研究で明確に示されました。今後は、具体的に脳の領域化のどの時期のどの分子機構がGroucho依存的制御を受けており、それぞれの分子機構がGrouchoの存在を介してどのように関わりを持っているのかを明らかにするために研究を継続します。間脳・中脳・神経堤・三叉神経節など多岐にわたる頭部神経系の変異それぞれに関して、その変異が起こる時期と領域の特定を行ない、またそれぞれの変異に関係する転写因子を単離することで、これら転写因子がどの特定変異に関与するか解析を行ないます。これらによって、脳のパターンをグローバルに制御する転写機構が明らかとなることが期待できます。またGrouchoは、その構造の中で少なくとも3カ所の転写因子結合部位を有し、相互作用する転写因子に応じて微妙な転写制御を行なうと考えられています。機能阻害に用いているペプチド配列は、このうちの一カ所に結合することが知られていますので、その他の部位での転写因子との結合を特異的に阻害することによって、全く新しい変異を導入できる可能性があります。
「頭部オーガナイザー」組織が頭部神経系をどのように部域化するかについても現段階では全く理解されていません。また、脳が自立的に領域化される機構についてもほとんど分かっていません。これら両者の理解が一体となって初めて脳の形づくりの理解が進むと考え、この両面からの解析を現在進行させています。
最後に、脳に代表される生き物の形を決めるしくみでは、非常に大まかな位置の情報がきめ細かな部域化を成立させることが分かって参りました(9)。この機構は、実は全く理解されておりません。ファジーな情報がシャープな情報へと変化するしくみ、これは生物が持つもっとも顕著な性質の一つでしょう。私たちは、頭部の形づくりの研究を通じてこのしくみに迫っていきたいと考えています。
参考文献
1) 時実利彦(1969)
目で見る脳=その構造と機能 東京大学出版会
2) Holley, S.A. (1997)
Fish are like flies are like frogs: conservation of dorsal-ventral patterning mechanisms. BioEssays 19: 281-284
3) Ahlberg, P.E. (1997)
How to keep a head in order. Nature 385: 489-490
4) Bally-Cuif, L. and Boncinelli, E. (1997)
Transcription factors and head formation in vertebrates. BioEssays, 19: 127-135
5) Koide, T., Umesono, K. and Hashimoto, C. (2002)
When does the anterior endomesoderm meet the anterior-most neuroectoderm during Xenopus gastrulation? Int. J. Dev. Biol. 46: 777-783
6) Fisher, A.L., Ohsako, S. and Caudy, M. (1996)
The WRPW motif of the Hairy-related basic Helix-Loop-Helix Repressor Proteins Acts as a 4-amino-acid transcription repression and protein-protein interaction domain. Mol. Cell. Biol.
16: 2670-2677
7) Zhu, C.C., Dyer, M.A., Uchikawa, M., Kondoh, H., Lagutin, O.V. and Oliver, G. (2002)
Six3-mediated auto repression and eye development requires its interaction with members of the Groucho-related family of co-repressors. Development 129: 2835-2849
8) Brickman, J.M., Micheal Jones, C., Clements, M., Smith, J.C. and Beddington, R.S.P. (2000)
Hex is a transcriptional repressor that contributes to anterior identity and suppresses Spemann organizer function. Development 127: 2303-2315
9) Holloway, D.M., Reinitz, J., Spirov, A. and Vanario-Alonso, C.E. (2002)
Sharp borders from fuzzy gradients. Trends Genet. 18: 385-387