

RESEARCH
腹側から見えた体の中心
脊椎動物の脊索はクモのどこ?
丸い卵に頭と足が生えたようなオオヒメグモの赤ちゃん。8本の足がきちんと体の両側にできるためには、丸い卵の左右を分ける中心軸が必要です。私たちには背骨がありますが、クモにはなにがあるのでしょう?
多様な形を持つ生きものを比較して形作りの共通点を探ろうと思ったら、どの種とどの種を比べたら知りたいことが一番よくわかるのか?これまで脊椎動物についての研究は、メダカやカエルやニワトリやマウス、そしてヒトで行われてきた。一方、背骨を持たない動物の代表は長い間ショウジョウバエだけだったが、実はショウジョウバエは、昆虫の中でもかなり特殊な発生をする生きものであり、脊椎動物の発生と比較するには困難な点が多いことがわかってきた。そこで私たちは背骨を持たない生きものの中からクモを選び、背骨を持つ生きものと持たない生きものの関係を見いだしたいと思っている。
1.クモの発生から脊索の起源を探る
私たち脊椎動物の背中と節足動物のお腹とが同一起源であるという考え方は、今から10年前に分子生物学の研究に基づく仮説として提唱され、多くの研究者の支持を集めた(図1)。しかし、その仮説では、脊椎動物の背中の発生に中心的な役割を果たす「脊索」が節足動物のどこに相当するのかについては全く説明ができていなかった。進化の過程で脊索は突然現れたのか?それとも、“もと”になった組織(または、領域)が初期の無脊椎動物にすでにあったのか? この問題は、脊椎動物が誕生するまでの経緯を理解する上で極めて重要である。最近、この問題を考える糸口が私たちのクモの研究から見えてきた。
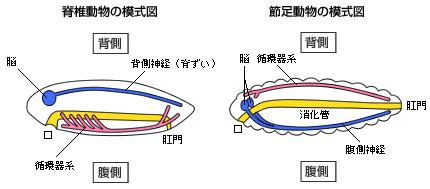
(図1) 脊椎動物と無脊椎動物の体の比較
脊椎動物の体では(左)、脳を含む中枢神経系が背中側にあり、腹側に消化管がある。これに対して節足動物は(右)、消化管の腹側に中枢神経がある。節足動物を上下逆さまにすると、脊椎動物と内部の諸器官の配置が一致することは19世紀から解剖学者や形態学者によって指摘されていた。さらに、近年の分子発生学から、発生過程で背腹の違いを生じさせる仕組みに脊椎動物とショウジョウバエで共通点があることが分かってきているが、これらのデータも脊椎動物の背と腹がショウジョウバエと逆であることを示唆している。
クモは発生の実験材料として有名ではないが、今から50年以上も前にホルムはイナズマクサグモの胚を用いた実験で、非常に面白い結果を報告している。クムルスと呼ばれる小さな領域を胚の反対側に移植することによって、お腹と背中を二組もつような“双子”の胚を作ったのである。このことは将来の背中とお腹の位置が卵の中で最初から厳密に決まっているのではなく、発生が始まってから細胞間の情報のやりとりを経て決まっていることを意味する。
2.背と腹を決める2つの分子DppとSog
私たちはdecapentaplegic (dpp) とshort gastrulation (sog) と呼ばれる2つの遺伝子に注目し、オオヒメグモの背中とお腹を決める仕組みを明らかにしてきた (註1)。dppとsogから作られるタンパク質は、いずれも細胞外に分泌される蛋白質(DppとSog)だ。Dppは細胞から細胞へ情報を伝えるシグナルとしての機能を果たし、一方Sogは、Dppに結合してそのシグナルを阻害する。まさに、細胞間の情報のやりとりを担う役者たちの代表例だ。
まずこれらの遺伝子の発現場所を見てみよう。受精30時間後の胚、将来の体を作る胚盤の中心が膨らんで、クムルスと呼ばれる領域が出現する。この時点では丸い卵のどちらが背側となるかはわからず、sogは赤道部よりも少し離れた胚盤の領域で同心円状に発現している(図2)。やがてクムルスは胚盤の周辺部に移動し、クムルスがたどり着いた側が背側となり、この反対側に将来の腹側が形成されるのである。この過程を通じてクムルスではdppが発現しており、sogの発現領域はそのクムルスとは反対側に狭められる。これらの発現は、クモの体作りにどのような意味を持つのだろうか。
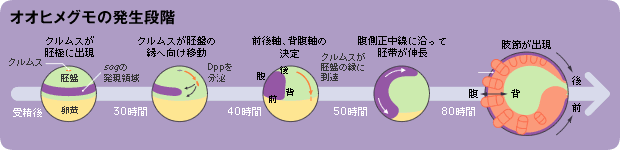
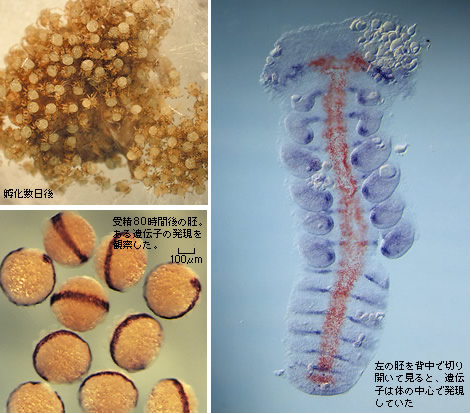
(図1) オオヒメグモの発生とsogの発現パターン
ある遺伝子が体作りにどうはたらいているかを調べるには、遺伝子を不活性化し、表現型がどう変わるかを見ればよい。クモの遺伝子を操作し人工的に変異体を作成することはできていないが、特定のmRNAを分解して遺伝子発現を抑えるRNA干渉法という方法をクモに応用することに成功した。これは、1本鎖RNA配列と結合する短い2本鎖RNAが細胞内に存在すると、そのRNAが選択的に分解される現象を利用したものである。2本鎖RNAを母グモに注入し、dppまたはsogがほとんど働いていない卵(初期胚)を得ることができた。その結果、dppが働かない状態では、背も腹もできず(図3)、一方、sogが働かない状態では、背はできるものの、腹側の中心となる構造ができなかった(図4)。つまり、本来腹になるべき領域がすべて背になってしまったのである。一連の実験結果をまとめると、オオヒメグモでは、まずdppの働きで背中の運命が決まり、次にsogの働きでお腹の運命が決まるのである。
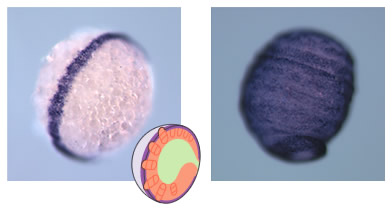
(図3) dppが働かなくなった胚
(左)腹側正中線を示すsogの発現とその模式図。
(右)実験でdppの発現を抑制すると、sogの発現は腹側に偏らず卵全体に広がってしまう。
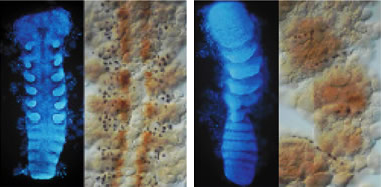
(図4) sogが働かなくなった胚
(左)正常胚を切開し、腹側から見たところ。全体像では対になった付属肢が観察できる。拡大図では、腹側神経節(紫色の細胞)を示す。
(右)sog遺伝子を働けなくすると、左右の付属肢が分離せず、神経節も正常に形成されなかった。
このようにsogがお腹の形成に働くことは、先に見た正常な胚のsogの発現パターンとも辻褄が合う。クムルスが分泌するDppがきっかけとなってsogの発現領域が腹側に狭められ、最終的には腹側正中線となり、その両脇には中枢神経が分化するのである。この腹側正中線と中枢神経が、sogの働きによって形成される腹側組織なのである(図5)。
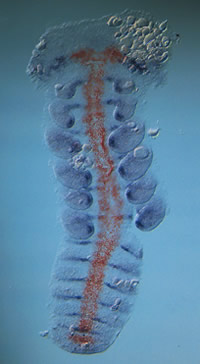
(図5)
赤色の染色がsog mRNAの発現、青色の染色はwnt16-like mRNAの発現で体節や付属肢を示す。sogの発現が腹側正中線に限局している様子がわかる。
(註1) 最新の結果は、以下の論文で発表した。
Akiyama-Oda, Y. and Oda, H. (2006) Axis specification in the spider embryo: dpp is required for radial-to-axial symmetry transformation and sog for ventral patterning. Development 133, 2347-2357.
この論文は以下のリンク先でpdfダウンロードできます。
論文はここからダウンロードできます。[PDF書類:5.1MB]
3.細胞の話し合いによって決まるクモの中心軸と脊椎動物の脊索
これらの結果から分かってきたことを簡潔に言えば、オオヒメグモではDppとSogを使って細胞間の“話し合い”をすることで腹側正中線が決まり、そこを中心に腹側のパターンができあがっている。このクモの仕組みは、脊椎動物が脊索を中心に背側のパターンを作る仕組みとよく似ている。クモ胚の腹側正中線も、脊椎動物胚の脊索も、胚の反対側から広がるDppシグナルを阻害する領域として形づくられ、付随して、隣接した領域に神経が誘導される。このようなクモと脊椎動物の類似が“他人のそら似”とは考えにくい(図6)。
私たちは今回の発見を基に、クモと脊椎動物の共通祖先について仮説を考えている。その仮説とは、「DppとSogによる細胞間の“話し合い”で体の中心軸(左右相称性の軸)を決めていた共通祖先がいたのではないか。そして、脊椎動物に向かった進化の過程では、その中心軸を脊索として体の内部に入れることに成功し、やがてその周りに骨が作られるようになったのではないか。」というものである。
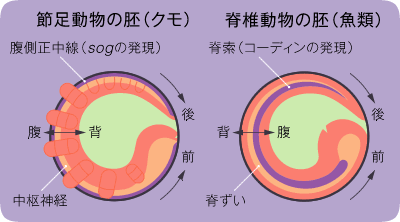
(図6) 節足動物と脊椎動物の形作りの比較
節足動物は体の外側(外胚葉)に、脊椎動物は体の内側(中胚葉)に中心軸を作る。sogとコーディン(chordin)は類似したタンパク質をコードしており、同じ起源を持つ遺伝子である。
4.ハエとクモの比較ではなく、クモとヒトの比較で見えたこと
これまで節足動物の知識は、優れたモデル生物であるショウジョウバエを中心に蓄積してきた。しかし、同じ節足動物でも発生や細胞のシステムには想像以上に違いがある(註2)。お腹を作る仕組みも、昆虫類のショウジョウバエと鋏角類のオオヒメグモでは大きく異なる。ショウジョウバエ胚の腹側領域の形成では、細胞間の“話し合い”ではなく、多核性胞胚で築かれるDorsalと呼ばれる核蛋白質の濃度勾配が中心的な役割を果たす。このようなショウジョウバエの仕組みはかなり特殊であり、脊椎動物と比較しても関連性を見いだすことはできない。
クモで得た結果から、私たちは、ショウジョウバエとクモの比較、そしてクモと脊椎動物の比較によって脊骨をもつ生きものと持たない生きものとの総合的な理解が得られるのではないかという期待を込めて、クモの研究に取り組んでいる。
(註2) ショウジョウバエとクモの発生の比較については、以下の記事を参照。
小田広樹(おだ・ひろき)
1996年京都大学大学院理学研究科博士課程修了。科学技術振興事業団月田細胞軸プロジェクトグループリーダー、JT生命誌研究館研究員を経て2006年より同主任研究員。
.jpg)

.jpg)
.jpg)










.png)