


TALK
形づくりが語る進化の物語
1.歴史をつくる
中村
生命誌のテーマはいつも動詞で考えています。「愛づる」で始まり「語る」「観る」「関わる」と続き、昨年はすべての始まりである「生る」を考えました。そこで、今年のテーマは「続く」です。生きものは「生る」の連続、つまり個体が生まれることのくり返しで38億年間続いてきました。「生きものが続く」と言ったとき、「遺伝子がつながっている」とされますが、これには具体性がありません。それにも関わらず、遺伝子が絶対で、それで決定されているような受け止め方がなされています。生殖細胞による受精と発生の過程を経て、個体が生まれるという形で性質が受け継がれ、その間に起きたDNAの変化によって、新しい種が生まれ、多様化していくわけです。ここに環境による選択も関わりますが、まずは個体発生プログラムの変化が進化の原動力だと思うのです。ここまでは読者のための説明で、倉谷さんには釈迦に説法、申し訳ありません。発生と進化をつなげた研究、最近では進化発生学という言葉も定着してきましたが、早くからそこに注目していらっしゃいましたよね。
倉谷
進化というと、上り階段のようなイメージが描かれがちですね。下等な動物から高等な動物に移行するかのように、上昇志向、或いは目的論で捉えられることが多いけれど、それは18世紀の考え方です。私が発生を通して進化を見た時に最も惹かれるのは、動物の体には変化しながらも変わらない部分があることなんです。それには胚の形が関わっていると思うんですよ。例えば、脊椎動物である限り避けて通れない胚の形が存在することは昔から知られていました。脊椎動物門ではこれを咽頭胚と呼びますが、他の動物門にもこのような胚の時期が存在するので、一般化してファイロタイプと呼びます。
19世紀前半にフランスのキュビエ(註1)が形態的特徴から大きな4つの動物群を分類した後、比較発生学者のフォン・ベーア(註2)が発生過程に注目し、胚の形にその4つの動物群の特徴を見出しました。そして、あるファイロタイプを保持した上での変化が多様な動物の形を作ると考えた。そして、ファイロタイプの異なる脊椎動物と節足動物は、形態的にも系統的にもまったくつながらないと考え、進化を否定しました。その背景にあるのはドイツ流の自然哲学というか、ある種の観念論(註3)です。僕たちが今、発生過程を観察しているとベーアの言う通りのことが見えるんだけれど、現代の科学に観念論は持ち込めませんから、それを裏付ける理論を考えなければいけない。そこで、発生プログラムでは胚の形に淘汰、或いは拘束がはたらいている可能性があると考えるわけです。
中村
同感です。それがどんな拘束か、それが知りたいですね。
倉谷
一種の構造的ネットワークを考えるとわかりやすいと思うんです。例えば、我々が使っているコンピュータのキーボードの左上には「QWERTY」と言う配列(註4)がありますが、これは本来はタイプライターのために開発されたものです。コンピュータを発明した時にキーボードを新しく作らずに、既にある配列を利用したわけです。実は、人間工学的には「QWERTY」よりも使い勝手のよい配列が考案されたこともありましたが、そのときすでにほとんどの人は「QWERTY」に慣れきっていた。そして社会のシステムも、それに基づいて固まってしまっていた。このように一旦決まった構造を根底から巻き返すことは事実上不可能で、既存のシステムの縛りを受けながら、せいぜい多少の改良を加えて、だましだまし使ってゆくしかない。遺伝コードも動物群によって多少の違いはありますが、原則は共通でしょう。
中村
生きものって38億年前に誕生した最初の生命体が持っていた細胞やDNAという枠組みを持ち続けている歴史的存在ですよね。だから生命誌です。過去を背負って生きる中で偶然の変化が起きてここまで来たわけで、ある目標に向かって変化したり形をつくったりしたことはないものですよね。
倉谷
そうなんです。でもその枠組みも絶対的ではないんじゃないかな。例えば、哺乳類が誕生する時には、脊椎動物の既存の枠組みから飛び出す変化がある。さまざまな器官を獲得する可能性を生むエポック・メイキングの瞬間があったはずです。
先日、私の研究所の外部評価委員から「カメ、ヤツメウナギ、マウスと、なぜそんなに色々な生きものを研究しているのか」と聞かれましたが、別に僕はヤツメウナギやマウスが好きで研究しているわけではなく、彼らを通して、動物でのエポック・メイキングな現象を探り、一つの動物群が生まれたきっかけを明らかにしたいのです。日本史なら大化の改新や明治維新に相当する、その後の展開を大きく変える一大事件です。哺乳類では、耳小骨(註5)が3つになった瞬間がその一つと考えられます。また、骨の形だけでなく化石に残らない胎盤や横隔膜の獲得も重要です。
中村
それを知るのに適した動物を見つけ出して調べると、ヤツメウナギでありカメであると。生命誌を見ていると「脊椎動物が生まれて5億年」とか簡単に言ってしまうのですが、今、耳小骨が3つになった瞬間とおっしゃった時の時間はどのくらいをイメージしていらしたのかしら。
倉谷
 自分でもどういう意味で瞬間という言葉を使ってしまったのかなと自問しているのですが、進化生物学者の頭の中で、何かまとまりのある分類群の枠が認識できた瞬間なのかもしれません。例えば、耳小骨が3つあることが、爬虫類から哺乳類を区別する一つの基準となった瞬間ですね。もっとも、それも細かく見てゆくと、とても瞬間などと言えた代物ではないのですが・・・。しかしそれでも、もし僕が哺乳類の祖先の誕生の頃に立ち会ったとしても、それが出現した当初は耳小骨が3つある、「一風変わった爬虫類」という程度にしか見なかったはずです。
自分でもどういう意味で瞬間という言葉を使ってしまったのかなと自問しているのですが、進化生物学者の頭の中で、何かまとまりのある分類群の枠が認識できた瞬間なのかもしれません。例えば、耳小骨が3つあることが、爬虫類から哺乳類を区別する一つの基準となった瞬間ですね。もっとも、それも細かく見てゆくと、とても瞬間などと言えた代物ではないのですが・・・。しかしそれでも、もし僕が哺乳類の祖先の誕生の頃に立ち会ったとしても、それが出現した当初は耳小骨が3つある、「一風変わった爬虫類」という程度にしか見なかったはずです。
中村
なるほど、人間が哺乳類が生まれたと認識した時が哺乳類誕生の瞬間というわけ。実際に起きたことというより、私たちの中での歴史ですね。
倉谷
認識論的にはそう言えるかもしれません。この3つの耳小骨の獲得が引き金となって、さまざまなニッチを獲得し、適応放散が起きてもはや爬虫類の一群とは呼べないほどの多様な種が生まれた。そこでこの多様な種を共通づけるものは何かと遡って考えると、3つの耳小骨となるのです。
中村
分類を歴史を語る作業と捉えるのは面白い。脊椎動物の大化の改新は3つの耳小骨と。
倉谷
結果論です。その当時はただの事件でも今から見るとその重要性がわかる。
中村
私たちがルネッサンスと呼ぶ時代にいた人たちは、「今がルネッサンスである」と思っていなかったわけで、そもそも歴史は後付けですね。起こったことの意味をきちんと考えて記述していくと初めて生命の歴史ができ上がる。
(註1) キュビエ【Georges Cuvier】
(1769―1832)
フランスの動物学者。比較解剖学の立場から、動物分類を秩序立て、動物の基本体制によって動物界を脊椎動物・関節動物・軟体動物・放射動物の4群に分けた。
(註2) フォン・ベーア【Karl Ernst von Bear】
(1792―1876)
ドイツの動物発生学者。ケーニヒスベルク大学教授。諸動物の発生を比較し、初期の胚が互いによく似ていることを指摘した。近代動物発生学の祖とされる。
(註3) ドイツ観念論
18世紀末から19世紀半ばにかけて発展したドイツ哲学の総称。カントに始まりヘーゲルによって完成される。自然に対し精神を優位とする立場をとり、世界を普遍的なイデア(理念)による体系として理解、構築する傾向を持つ。
(註4) QWERTY配列
1882年に考案された英文タイプライターのキー配列の一つ。アルファベットを打つキーが左上から「QWERTY・・・」であるためこの名で呼ばれる。
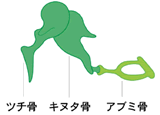
(註5) 耳小骨
中耳の中にある小骨。鼓膜の振動を内耳に伝える。ほとんどの動物はアブミ骨のみを持つが、哺乳類だけがアブミ骨、キヌタ骨、ツチ骨の3つの耳小骨を持つ。
2.分類から進化をよみとく
倉谷
脊椎動物は大きくは顎の有無で分類されますが、分子系統樹を描くと、顎口類と円口類の分岐点付近では、顎を持たない顎口類もかなり存在します(図1)。つまり、顎口類は、顎を獲得することによって誕生したとは言えない。私は、本来1つだった鼻の孔が2つになることが顎をつくる下地となったと考えています。偶然、鼻の孔が2つのものができ、それが顎の形態的進化を可能にして、現在のような顎口類の形になったのではないか。
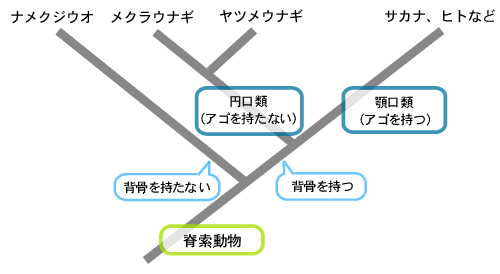
(図1)
円口類説に基づく脊椎動物の系統樹
倉谷滋『動物進化形態学』(東京大学出版2004)より改変
中村
では、脊椎動物の始まりは顎の誕生に先立つ2つの鼻の孔の獲得と考えていいのですか。
倉谷
まだそうは決められないんです。顎より背骨が先だったかもしれませんし、神経提細胞や間葉細胞が発生過程で現れることが大きかったかもしれないし。今、そこを追うためにホヤやナメクジウオのような脊索動物を勉強しているのですが。その幼生のボディプランを見ると、背側に神経管、真ん中に脊索、腹側に腸管が通って、両側に中胚葉または体腔があるという点では、脊椎動物の基本体制と共通しているんです。ただ、形ができる時のサイズがまったく違う。
中村
サイズということは、細胞の数ですか?
倉谷
ええ、脊椎動物と脊索動物とでは、胚の形を組み上げるときの細胞の数がえらく違うんです。分子レベルでも、シグナル分子が拡散するべき距離が変化すれば、さまざまなメカニズムが変化せざるを得ないでしょう。比較形態学はパターンだけを扱ってきましたが、分子生物学が入ってきて体の大きさとか細胞の数も考えなければいけなくなりました。私が最も気になるのは間葉細胞の量です。そこからできた骨を基本にさらに大きな変化が起こったはず。
中村
色々な器官をつくり得る細胞の数が増え、可能性が広がったということ。
倉谷
細胞の数が増えることが質的向上につながると考えられます。細胞が増えれば、それぞれの細胞の中での遺伝子のはたらき方に変化が生まれる可能性がある。シグナル・カスケード(註6)の変化によってそれまでとは異なる分化のシナリオが誕生するかもしれないでしょう。どういう過程を経て今のような生きものが生まれたのかはまだまだわかりませんが、これ面白い問題だと思っています。
中村
量から質に変わるところを知るには、やはりそこではたらく遺伝子、全体としてはゲノムを見ていくことになるわけで、分類から進化という時間を見ていく一つの切り口ですね。ただ、ゲノムへ行く時に形の問題として考えることもありますでしょ。
倉谷
 もちろんです。最初に話した哺乳類の耳小骨の問題は昔から興味があって長々とやっている仕事ですが、生きものの形づくりを考えるためのゲーテ(註7)やジョフロア(註8)、あるいはオーウェン(註9)といった学者たちによる初期の形態学では、まずエラや椎骨のつながった原型が描かれ、次に各部分がそれぞれの場所において形を変えてさまざまな機能を持ち、顎や耳が生まれるとされました。エラからの変化の基礎問題が顎の誕生ならば、哺乳類の耳小骨の誕生は応用問題と言える。まず顎ができ、その関節部が内側に移動して音響装置としてのはたらきを持った。それが耳小骨です。
もちろんです。最初に話した哺乳類の耳小骨の問題は昔から興味があって長々とやっている仕事ですが、生きものの形づくりを考えるためのゲーテ(註7)やジョフロア(註8)、あるいはオーウェン(註9)といった学者たちによる初期の形態学では、まずエラや椎骨のつながった原型が描かれ、次に各部分がそれぞれの場所において形を変えてさまざまな機能を持ち、顎や耳が生まれるとされました。エラからの変化の基礎問題が顎の誕生ならば、哺乳類の耳小骨の誕生は応用問題と言える。まず顎ができ、その関節部が内側に移動して音響装置としてのはたらきを持った。それが耳小骨です。
自分自身が哺乳類だから、哺乳類を決めることになったこの問題に惹かれているのですが、それとは別に、ここには進化における機能的変化と物理的変化に対応関係をつけられる可能性を感じているんです。耳小骨の一つひとつが、魚のエラのどの部分にあたるかが言える。原型は変化せずに、機能がまったく変わっているわけです。相対的位置関係は変えずに、形状を変化させ、物理的な機能を変化させたことが重要なのです。劇的な転換をもたらしたこの問題を「なぜ」と問うのは非常に難しく、だからこそ挑戦しているのですが、その面白さの在りかをうまく説明できないんですよ。
中村
相同器官の例を枚挙するだけでは、進化は語れないし。
倉谷
いわゆるツールキット遺伝子(註10)の発見を目標とするなら、脊椎動物の顎やエラの相同性を証明するだけで研究は達成されます。我々が知りたいのは、なぜ脊椎動物は発生パターンの基本的な形態を同じくしながら、これほど多様な形を生みだしたのか、その変化が種ごとのまとまりを作っているけれど、どういう経緯でそれを獲得したのかということです。
これは、私を育ててくれた比較形態学が19世紀以来取り組んできた金字塔のような問題なんです。彼らには見えていなかったゲノムを視野に入れると、発生パターンの過激な進化が見えてきて驚きます。
中村
哺乳類に限らずフクロウのような鳥類にも耳はありますよね。その中で哺乳類の耳は特殊なのですか。
倉谷
基本的には同じで、3つの耳小骨のうち、内耳に統合しているアブミ骨という骨は鳥類にもあります。ところが、残りの2つの骨と、おそらく鼓膜は哺乳類にしかありません。鼓膜の由来に関してはさまざまな説がありますが、私は哺乳類になる時に新しく獲得したものだと思っています。もちろんトリにもちゃんと音は聞こえています。鳴き声でコミュニケーションしますし、人の物真似もするぐらいですからね(笑)。なぜ哺乳類は、トリとは異なった方法で中耳を作らねばならなかったのか、そしてどういういきさつでそれを獲得したのか。知りたいところです。
脊椎動物は、非常に複雑な体の構造をしているけれど、パズル遊びのようなところがあって、よく見るとさまざまな動物と同じ筋肉や骨を使っています。コウモリの翼と僕らの腕は相同で、上腕骨が伸びて皮膜が張ったというのはよく知られていますよね。ではなぜそうなったのか。進化の「なぜ」を目的論的に問いかけても難しいですね。
中村
そもそも、科学は「何がいかに」を語るもので、「なぜ」に対する答えはしない約束でしょ。
倉谷
生きものの形態は、環境に合わせて上手く機能しているから、初めから適応を目指して設計されたかのように見えるけど、それを導いたのは自然淘汰だということですよね。だからこそ、その経緯を知ることが楽しい。種が多様化するためには、ある順番が必要ですし、新しい個体がつくられるためには、祖先が利用してきた発生の方法を利用するしかない。裏を返せば、そうした縛りがあるから、分類学的な入れ子関係や類縁関係がはっきり示せるのでしょう。
中村
最初に、形はさまざまだけれど、どこかに変わらないものがあるということが見たいとおっしゃった、そのことですね。
倉谷
ある目的のもとにゼロから系統を作ったら、歴史を背負った関係などないので、分類はできなくなる。分類学が成立するということ自体が進化の証だというのが私の持論です。
(註6) シグナル・カスケード【signal cascade】
細胞外の刺激が細胞内に伝えられる時、複数のタンパク質が連鎖反応的に信号を送る仕組み。カスケードは滝の意。
(註7) ゲーテ【Johann Wolfgang von Goethe】
(1749-1832)
ドイツの詩人、作家。比較解剖学の研究者としても有名で、「形態学」という用語を最初に使い始めたとされる。
(註8) ジョフロア【Geoffroy Saint-Hilare, Etienne】
(1772-1844)
フランスの博物学者・解剖学者。相同性を定義した。近代的奇形学の創始者と言われる。
(註9) オーウェン【Richard Owen】
(1804-1892)
イギリスの比較形態学者。大英博物館自然史館の初代館長。キュビエの影響を受け、比較解剖学に相同と相似の概念を導入した。
(註10) ツールキット遺伝子
形態形成を制御する一群の遺伝子で、多くは転写因子とシグナル分子からなり、他の遺伝子の発現を調節する。動物門を越えて保存されており、ホメオボックス遺伝子もその一つ。
3.胚の選択が導く形づくり
中村
これまでのお話、その通りだと思うのですが、そこでさて進化の物語をどう書くか。そこが難しいですね。
倉谷
いつも考えているのですが、なかなか答えは出ません。ゲノムにこだわる研究者は、ゲノムのはたらきから動物それぞれの特徴を描くことを目標にしていると思いますが、私の場合はもともと形態学者なので、形態学的な特徴は理解しています。その上で、なぜゲノムを研究しているかというと、脊椎動物の胚の形ができるところでのゲノムのはたらきが気になるのです。
中村
同じように見える胚から、マウスが生まれたり鳥が生まれたり。そこではたらくのは、マウスのゲノムであり、鳥のゲノムですからね。胚の時期に起こる淘汰が進化につながる、つまり胚は個体としての始まりであると同時に、そこに起きる変化が種の始まりとも言えるわけで、変わりながら続くことの鍵は胚ということになりますね。ここで個体発生と系統発生とが関わってくる。
倉谷
ヘッケル(註11)に始まるいわゆる反復説は発生過程と進化のプロセスの間に平行関係を見出し、以来、多くの研究者が発生に進化をなぞらえて反復説を証明しようとしましたが、成功しませんでした。むしろ最近では、それが不正確であるということだけが強調されている。ただ、それでも胚発生過程が進化を可能にしていることだけは間違いないと思うのです。
中村
私もそう思います。
倉谷
胚の選択か、成体の選択か。進化を考える上で非常に難しいところですけどね。先ほど、2階ギャラリーの展示(註12)で、右巻と左巻のマイマイを見せていただきましたが、連続的な変化をベースにして成体で巻の向きを逆転するためには、一旦殻を棒のように伸ばしてから再び巻き直さなければならない。でも、発生のヒエラルキーの中で、ごく初期に身体の左右性に関わるような遺伝子に変異が起これば、以降の体づくりのプログラムをすべて逆向きに変更できます。
中村
成体の選択は、大きさなどであって、種の多様化を生み出したのは、やはり胚の選択だと思う方が考えやすいですね。
倉谷
先ほどの「鼻の孔が2つ」ということも発生上の制約なんですよ。言葉で説明するのは難しいのですが、鼻腔のしきりである軟骨の形成と顎の誕生には関係があり、鼻の孔が1つだと、軟骨を作る空間がなく、顎の形成に進めないのです。
中村
軟骨ができてから、鼻の孔ができたというわけではないのね。
倉谷
整合性からすると軟骨があって鼻の孔が1つある状態は難しい。発生プロセスの途中で鼻の孔を2つ持つ必要があったのだと思います。
中村
顎のような器官が生まれるためには、さまざまな可能性に満ちた発生過程で、色々な道を試しながら、現在の有顎類の発生パターンに落ち着いたのでしょうね。
倉谷
大人の形を選択することによって、その形を導く発生プログラムを選択しているわけです。選択によっては、胚発生そのものが全うできない場合もあるでしょう。
中村
 形、分類、進化、発生。生きものについて考える時、これらは皆重要でしかも関連していますね。遺伝子のはたらきは大事だけれど、それだけで何かを語ろうとしても無理。最近形態学や分類学はそれを勉強しませんでしょ。倉谷さんはそこができていらっしゃるし、研究の歴史も勉強しているので強いですね。
形、分類、進化、発生。生きものについて考える時、これらは皆重要でしかも関連していますね。遺伝子のはたらきは大事だけれど、それだけで何かを語ろうとしても無理。最近形態学や分類学はそれを勉強しませんでしょ。倉谷さんはそこができていらっしゃるし、研究の歴史も勉強しているので強いですね。
「骨と形」の展示(註13)の制作で、倉谷さんにエラから顎ができると教えていただいてびっくり。無顎類と有顎類については、高校の生物学で習いましたが、ただ暗記だったので面白くなかった。イタリア語から翻訳された進化に関する本の内容のチェックを頼まれた時、顎のない魚は口を開いて流れ込んでくるものを飲み込むしかないが、顎ができたら自分でパクリと食べることができると書いてあったのです。顎って生き方を変えるほどの意味を持つんだと驚いたのです。形、分類、進化が生き方にもつながるわけですね。
倉谷
その通りなんですが、そこで顎が誕生すると無顎類は一掃されるかというとそうではなく、無顎類と有顎類の共存期間はかなり長いんですよ。
中村
そこが生きものの面白いところですね。顎を使って積極的に餌を食べられることはすごいけれど、それだけが生き方ではなく、口を開いているだけでも餌は充分入ってくる条件下ではその生き方で充分というわけですね。
倉谷
しかし、あるナマズの仲間は、せっかく獲得した顎を吸盤のように用いて岩に吸い付いたり苔を削ぎ落として食べたり、古生代の無顎類と似たような暮らしをしています。それにその時期には、顎だけでなく、鰭を獲得したとか、感覚器の充実、免疫系や消化機能などの生理機能の増大など色々な機能が変化している可能性があるのです。
中村
エポック・メイキングな時期はある一つの機能の変化で代表されるのではなく、さまざまな機能が関わり合って変化する時なんですね。なるほど。機械はバージョンアップに伴い古い機能は捨てられますが、生きものの場合はバージョンアップしても、それ以前のものも捨てずに残しますでしょ。また古いものが同じところで競争したら負けるかもしれないけれど、棲み分けして全体として上手に共存することができる。
倉谷
そうですね、現在の無顎類のヤツメウナギとヌタウナギは、動物の死骸をあさったり、寄生したりという独特の生活型を獲得し、そうした特殊化によって生き延びている。同じニッチを占める生きもの同士なら効率の悪い方は負けてしまいますけれど。
中村
そのようにして多様になっているのが生きものの面白さですね。
(註11) ヘッケル【Ernst Heinrich Hckel】
(1834-1919)
ドイツの動物学者・思想家。ダーウィンの進化論に基づいて、個体発生は種の系統発生の短縮されたものであるという反復説を提出し、ベーアとは異なり、ある動物の初期の胚はそれより下等な動物の成体の形態に相当するとした。

(註12) 愛づる・時 ~生命誌がひらく生きものの絵巻物語~
2004年4月より展示中。
ヒダリマキマイマイ(左)からごく最近分岐したアオモリマイマイ(右)。
共通祖先は右巻で、左巻に分岐したのち、先祖返りのように右巻に分岐。
(註13) 骨と形~骨ってこんなに変わるもの?
2003年4月より展示中。
4.「いつ、どこで、どのように」が鍵
中村
これまでのお話のような考え方を確実なものにしていくために、今どんなことを?
倉谷
色々ありますが、例えばカメの甲羅は面白いですよ。「骨と形」展の甲羅を開いた標本は素晴らしい(図2)。背骨と肋骨が1つの塊になって甲羅の背側半分を作っていることが一目でわかりますね。ところが、なぜそうなったかとなると謎なのです。
僕らの肩甲骨は胸郭の外側についていますが、カメの肩甲骨は逆に甲羅の内側にあるんです。先ほどから話題にしてきた形の縛りからすると、この逸脱は非常に都合の悪い例ですね。基本的な形の共通性が維持されていなければ、分類はできません。ところがよく観察すると、カメの肩甲骨も甲羅の外側にある時期があるんです。
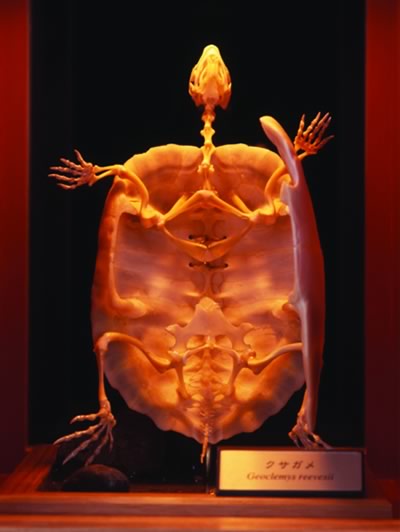
(図2)
クサガメの骨格標本。「骨と形」展より。
中村
発生の途中で?
倉谷
はい。実はカメの肋骨は短いので、その隙間に肩甲骨が潜り込めるのです。肋骨の基本パターンはカメもトカゲも我々も同じですが、カメの肋骨だけは体壁の中に伸びず、それに沿って横に伸びていくんです。調べてみたら、甲稜と呼ばれるカメの肋骨の先端部で発現する遺伝子をいくつか発見しました。
ところが、それはカメに特有の遺伝子ではなく、ニワトリにも我々にもマウスにもある遺伝子なんです。それをカメは独特の使い方をしている。新しい遺伝子の獲得ではなく、使い方が問題だということがここではっきりしたわけです。ただ、何が遺伝子を発現させる引き金かはまだわからない。
中村
研究館の宮田顧問も、カンブリアの大爆発が起こる以前に、多様化に関わる遺伝子セットはすべて用意されていたことを示し、遺伝子の問題は、いつ、どのように使われるのかというソフトの問題だと考えて仕事を進めています。生物学では、物理学のような法則は見つかりません。結局、遺伝子の使い方のようなソフトのデータをきちんと整理して、そこにあるきまりを明らかにしていくことになるんでしょうね。
倉谷
カメだけに発現する遺伝子は、他の動物の遺伝子ネットワークによく顔を出す常連さんです。生きものの歴史と同じように、研究にもエポック・メイキングと言えるものがあって、ある遺伝子の発現がわかったことで、芋づる式に次々とさまざまな遺伝子が発見されることがありますね。Wntシグナル経路(canonical wnt signaling pathway)に登場する分子群などその例ですね。遺伝子カスケードやシグナル経路は既知のもので、それがどこで機能しているかを発見することで新たな役割が見えてくる。
中村
その場所でその遺伝子を使うに到った経路が知りたいですね。
倉谷
ええ。例えばカメに特有の遺伝子発現機構を見るとしたら、今の研究の流れで行けば、発見した遺伝子を調節する遺伝子を探ることになるんでしょうけれど、私は細胞と細胞の出会いによる相互作用を考えたいんです。それにカメ特有のものはないだろうかと。
中村
私は化学から分子生物学に入りましたから、発生学は自分で実験をしたことがありませんので、周りの人から教えてもらってなんとか考え方を身につけたいと思うのですが、とても単純な疑問で、脊椎動物は皆、背骨や肋骨を持つ仲間で最終的にはおさまるところにおさまるのに発生過程が異なるでしょう。カエルもニワトリも、基本は同じにしてくれればよいのに、なぜ意地悪みたいに手順を逆転させたりするのかしらと、分子生物学的な思考では思ってしまうんです。
倉谷
2種類の生きものに相同な遺伝子が発見された場合、その遺伝子は共通祖先に由来していると言えますが、相同な器官が違う遺伝子でできているのを説明するのは難しい。しかもそういう例はたくさんあるんですよ。
中村
同じ祖先から受け継いだものを使いまわしているのに、使いまわし方が気まぐれというか。使い方に幅が許容されている。枠組みとして決めるところと許容の兼ね合いでできてくるんですね。
倉谷
今日のテーマに則すなら、「形の続き方」が1つの鍵ですね。動物群を定義づけるボディプラン(註14)はある。これはめったなことでは外せないと。しかしそれが少しずつ変化しながら分岐を重ねて放散していくと、連続的な形の変化が生まれ、そこに歴史的な道筋が見えるんです。ゲノムの進化は、同じ系統の中では重複や欠失が起きる程度で、ほぼ連続的に変化していきます。ところが、形態の進化とゲノムの進化が対応しているかというと、そこが違うんですよ。
中村
まさにそこです。そこがポイント。従来の科学の思考回路で考えると混乱します。生きものをよく見て、生きものを考える回路を作らなければいけないと思っているんです。
倉谷
 昔の反復説は、発生中の胚の形が進化の過程をなぞっていると考えました。ある系統の中での遺伝子の進化と形態の進化の対応を仮定して、常に同じ遺伝子が同じ形に結びつくと考えたがっているのが現代の反復説だと思います。でもそうじゃない。ゲノムの持つ拘束と形の淘汰に関係があることは確かですが、そこには動物ごとに相当に異なるロジックがあると思います。
昔の反復説は、発生中の胚の形が進化の過程をなぞっていると考えました。ある系統の中での遺伝子の進化と形態の進化の対応を仮定して、常に同じ遺伝子が同じ形に結びつくと考えたがっているのが現代の反復説だと思います。でもそうじゃない。ゲノムの持つ拘束と形の淘汰に関係があることは確かですが、そこには動物ごとに相当に異なるロジックがあると思います。
中村
その関係が合理的に説明できませんね。たまたま思いつきでやりましたみたいなところがたくさんありますし。
倉谷
目という器官はすべての後生動物(註15)が持つものですし、目の形態形成に関わるマスター制御遺伝子Pax-6(註16)も、ほとんどの動物が持つ普遍的な遺伝子だということがわかりました。形態学者は目のない変異体に出会った時、網膜や、虹彩、レンズや、視神経や、外眼筋といった、目に付随する器官を枚挙するから、とても大きな変化が起きたように感じますが、ゲノムのレベルではPax-6という遺伝子一つの変化でしかない。遺伝子と形を対応させるのが如何に難しいかを教えてくれる例です。
中村
しかも動物によって目の形、そこで用いる分子が違うわけで。Pax-6という遺伝子は共通でも、発生というカスケードの結果、形態的にはまったく違う目ができてきますよね。
倉谷
そう、遺伝子は単独で等しい重みを持つのではなく、ネットワークの中で各々の重要性を持っている。そうした遺伝子の重要性が淘汰を通してどのように保守され、今発現しているのかを認識しなければいけません。
ゲノムから見る場合、完全に分析主義的ですから、形態学の視点とゲノムから見えてくることを一対一で対応させるのは無理です。形態学者の立場からすると、形態学と還元主義はなじまないし、極端な細分化には危険を感じます。そこでもっと発生を見たらどうだろうと思うわけです。
中村
形態学とゲノムと発生をつなぐことの重要性は、今声を大きくして指摘すべきことですね。
倉谷
あらゆる段階の発生を見て、特定の発生プロセスが形づくりに関与しているのか、どの遺伝子がどんな決定的な重要性を持っているのかを探れば、形態と発生プロセスはつながるはずです。
中村
比較形態学も進化学も、一般的にはでき上がった個体を対象にしますが、実はでき上がる過程が大事。あらゆることを考える鍵ですね。
(註14) ボディプラン
動物の体の基本的な体制。分類学ではボディプランが同じ動物を「門」に分類する。
(註15) 後生動物
多細胞体制をもつ動物の総称で、ヘッケルが単細胞性の原生生物と対置するものとして造語。生物五界説における動物界と同じで、海綿動物から脊椎動物にわたる。
(註16) Pax-6
1994年に、ショウジョウバエで発見された目の発生のマスター制御遺伝子。Pax-6を本来の発現場所ではないところではたらかせることにより、例えば脚の先に目をつくらせることができる。
※関連記事 生命誌ジャーナル12号「ゲーリング博士が語る 目の進化の物語:ウォルター・ゲーリング」
5.ネットワークは相互作用
倉谷
以前に阿形清和(註17)さんと話したのですが、例えばカンブリア紀にいたアノマロカリス(註18)っておかしな生きものがいますでしょ。あれを作ろうしたら、どうします?
中村
現在の生きもので似ているところを参考にしてつくりますね。
倉谷
そう、既に知っている形態形成機構を利用しようとしますね。その時、ゲノムが何通りあるのだろうと考えました。ゲノムは決定論的に形態を決めるのだろうか、多少の遺伝子の位置の変化があるとしたらゲノムの縛りとボディプランの関係はどうなるのか、そうしたことを考えるよい思考実験になります。
中村
なるほど、おもしろいですね。具体的にはどんな生きものから出発すればよいのでしょう。
倉谷
現存の生きものではカギムシが近いと思いますが、カギムシのゲノムを見るのはルール違反ということにして(笑)、ゼロから考えます。アノマロカリスゲノムはどんな形になるのだろうか。これはゲノムを見てボディプランに言及できるかどうかが、本当にゲノムから形を説明できるのかという問いへの試金石になるでしょうね。実現はずっと先の話だと思いますが。
中村
おもしろい。パっと見て、これはカエルゲノム、これはハエゲノムと特定の動物がわかるようになるかしら。
倉谷
コーディング領域(註19)なら、慣れている人は特定の分子でわかるでしょうが、それはゲノム全体をわかるというのとは大分違う気がします。
中村
 ゲノムは塩基配列としては一次元に並んでいますけれど、現実に細胞の中ではたらくときは違う姿になりますね。ネットワークではたらくゲノムがイメージができたらいいなと思うのですが。
ゲノムは塩基配列としては一次元に並んでいますけれど、現実に細胞の中ではたらくときは違う姿になりますね。ネットワークではたらくゲノムがイメージができたらいいなと思うのですが。
倉谷
私はできません(笑)。実は、ネットワークという呼び方はシステマティックな印象があって、抵抗を感じているんです。私のイメージでは、非常に入り組んだ網の目の中に、いつのまにか中枢機関というか、何か「ハブ」のようなものが成立し、その近くにマスターコントロール遺伝子が次々に成立してくるような風景ですね。
中村
酵母の全タンパク質を示した図(註20)はまさにインターネットですよ。ハブもあって。
倉谷
生物とネットの進化はよく似ていて、インターネットの提唱者が今もインターネットを支配しているかというとそんなことはない。新たな重要性を帯びたハブを基準にシステムが追随し、さらに分岐して多様化していくイメージです。
中村
機械の回線と違って、ゲノムの回路は特定の機能のために部品を組み立てるのではありません。ネットワークのつくり方は種ごとに異なるのだから、どこから手をつけていいのか。
倉谷
ある程度データが集まっているモデル生物を使って、少しでも複雑さを軽減しようとしている研究者は多く、私の研究所でも自分以外のラボはみなそうですね。
中村
でも、モデル動物だけを見ていても、複雑さは理解できないでしょう。先ほどおっしゃったエポック・メイキングなところについても、いくつかの種を見比べて初めてわかるわけでしょう。
倉谷
複雑さは、順列の組み合わせに由来する幾何級数的な爆発から来るわけですね。要素自体は有限個ですし、細胞の数も自然数です。発生は、ニワトリならばせいぜい20日間で終了するプロセスです。人間の賢さをもってすれば理解できるだろうと、アリストテレス以来、皆でそれを追いかけてきたんだけれど。
中村
その通りですね。始まりは一つの細胞で、総数は有限個であることはわかっているのだから、スーパーコンピュータを使えば20日間くらい追いかけられそうな気もしますし、一方でいやいやそんなことでは・・・という気持ちもありますから・・・。
倉谷
神戸の理研では2000年から取り組んでいますが、その1日分も完成していないかもしれない。私の専門は脊椎動物ですが、ショウジョウバエの研究者に「それだけ多くの世界中の科学者が日夜しのぎを削って研究しているのだから、君たちはショウジョウバエくらい、もう作れるんじゃないか」と冗談を言ったことがあります。ショウジョウバエのような小さな生きものすら理解できていないのです。
中村
今はスーパーコンピュータが入りますね。シミュレーションでも無理かしら。
倉谷
セル・オートマトン(註21)として、自己複製する細胞をつくろうとすると、最初の一つの細胞をつくるだけでも月面くらいの面積を持ったコンピュータのモニターが必要だと計算した人がいましたよ。
中村
それは現実味がありません。
倉谷
量から見るとヒトゲノムなんて単純です。あるゲノム学者の表現を借りると、1m3の立方体を作って1mmに刻み目を入れて、格子線を張ると、その交点に塩基を1個ずつ置いていくと、32億塩基が1m3の立方体、僕らがおふろに入る時の水の量より少し多いぐらいの容積におさまってしまう。
発生過程や遺伝子ネットワークの複雑さは桁が違うでしょう。ヒトゲノムの塩基配列を文字にしたものは、ちょっと厚目の本におさまりますが、そこから情報を読み出してつくられた自分の体を、電子顕微鏡で観察して記述しようものなら、とても一冊の本では済みません。ゲノムの複雑さを塩基配列の情報量として認識してはいけないと思います。
中村
でも、現実に一次元情報としての塩基配列は存在していますよね。一次元情報にすべてをおさめたわけで、生きものって変なやつだと思います。
倉谷
 一次元情報というのは曲者ですよ。私は音楽が好きで、例えばバッハを聞いて感動したりしますが、記憶の中の音楽は、演目や楽器のパートやメロディに分割されている。1つのスピーカーから流れてくる音を聞くときも同じことで、我々の聴覚機能は非常に多元的で、そういった要素の質的部分を感知し、目を閉じていても演奏したり、唄ったりしている人たちの位置とか、振る舞いを想像することすらできる。ところが、レコードやCDは一次元情報を記述するものでしかないし、その中に楽器ごとの情報が分かれて入っているわけではない。ですから、DNAという一次元情報が生物の三次元的身体を作ったとしても、それ自体は今さら驚くことではありませんよね。
一次元情報というのは曲者ですよ。私は音楽が好きで、例えばバッハを聞いて感動したりしますが、記憶の中の音楽は、演目や楽器のパートやメロディに分割されている。1つのスピーカーから流れてくる音を聞くときも同じことで、我々の聴覚機能は非常に多元的で、そういった要素の質的部分を感知し、目を閉じていても演奏したり、唄ったりしている人たちの位置とか、振る舞いを想像することすらできる。ところが、レコードやCDは一次元情報を記述するものでしかないし、その中に楽器ごとの情報が分かれて入っているわけではない。ですから、DNAという一次元情報が生物の三次元的身体を作ったとしても、それ自体は今さら驚くことではありませんよね。
中村
その例えはおもしろいですね。確かに音楽には時間がある。ゲノムを楽譜とした演奏。なぜゲノムは一次元なのかずっと不思議だったのですが、そういえば文字も一次元ですよね。
倉谷
本を読んで泣く人の一体どこに感動があるのかと言えば、それはページの上の一次元の文字列の中にはないですよね。情報処理の過程で生まれる何らかの相互作用だと思います。文字も音も、人間が記憶媒体に使っているものは全部一次元というのは、偶然ではない気がします。
中村
言われてみれば、生きものの情報処理がそもそも一次元で行われている。脳を研究されている方は、聴覚が一番基本的な感覚だとおっしゃいます。
(註17) 阿形清和
現京都大学教授。
(註18) アノマロカリス【Anomalocaris】
カンブリア紀中期(約5億2500万年~5億500万年前)の海に生息していた捕食性動物。現生のカギムシ(有爪動物門)に近縁と推定されている。
(註19) コーディング領域【coding sequence】
ゲノムの遺伝子領域でタンパク質のアミノ酸配列を規定する配列。
(註20) 関連記事
(註21) セル・オートマトン【Cellular automaton】
アメリカの数学者ノイマン(1903-1957)によって考案された自己増殖系の計算モデル。平面に広がる格子状のセルで構成され、各セルは自己と近傍の状態のみに従って内部構造を変化させ、系全体の動きを定める。
6.反復はおもしろい
中村
 倉谷さんは「個体発生は進化を繰り返す」というヘッケルの反復説は立証されていないことを強調なさいますね。それはわかるのですが、でも反復という概念はとても魅力的です。生物の体にはよく反復構造が現れますし、ゲノムの中にも反復配列はたくさんあります。こういうことがまだわかっていない時代のヘッケルは反復をどう捉えていたのでしょうか。
倉谷さんは「個体発生は進化を繰り返す」というヘッケルの反復説は立証されていないことを強調なさいますね。それはわかるのですが、でも反復という概念はとても魅力的です。生物の体にはよく反復構造が現れますし、ゲノムの中にも反復配列はたくさんあります。こういうことがまだわかっていない時代のヘッケルは反復をどう捉えていたのでしょうか。
倉谷
ヘッケルは、一種のロマン主義のもと、自然の壮大な摂理を見出そうと、彼なりの進化論的原型論を思い描いたのだと思います。この世のすべてをイデア(註22)で理解することによって、あらゆる変化やパターンの多様性を整合的に説明できると考えた。脊椎動物を統一的に理解しようと、胚の形に、哺乳類のイデア、霊長類のイデア、そして人のイデアというように、細分化したイデアの繰り込み構造を見出すのです。これは私の考えですが、原型論が反復説を後押ししたことは間違いありません。
中村
多様性は重要だけれど、その背後に普遍性を見ようとする努力はずっと行われてきて、今も続いているわけですね。
倉谷
生物学をやるには、まず分類学を身につけろと教えられましたが、それは間違っていないと思います。分類学はこの世のカタログの見方を教える基礎教育です。創造説が優位だった18世紀、リンネ(註23)は神の英知を探るために、自然のシステムがどのように構築されているか、階層ごとの支配関係を理解しようとして、システム化された自然の認識方法をつくろうとした。現代で言えば、化石を含むあらゆる動物の系統関係を明らかにしなければ、進化のプロセスは理解できないという考えに近い。いわばヘッケルは、それを発生プロセスにも応用しようとしたのではないでしょうか。
私にも分類によって物事を系統的に整理して見る癖があります。本来発生プロセスに名前はついていません。「受精卵」「2細胞期」「桑実胚」あたりは目で見た細胞の特徴を当てはめていますが、「神経胚」には、いつからいつまでが神経胚かという明確な線引きはありません。胚発生を分類しようとすると、人間が考えた記号を当てはめざるをえない。反復説は、脊椎動物のファイロタイプ以降に「顎口類期」、「サメ期」、「霊長類期」と進化の段階を割り当てたもので、自然の体系化です。発生プロセスの認識としては、際どいところだと思います。
中村
人間は分類したり、何かを何かに当てはめるのが好きで、そうすると安心して納得しますでしょ。何とかそうしたいところだけれど、生きものって難しいですね。
倉谷
 我々が発生にそうしたシステムを見てしまうのには2つの可能性があると思います。1つは、実際に生きものが我々が感知したシステムどおりの序列で進化を遂げており、それを見てシステムと呼んでいるという可能性です。これは、分類学や系統学に関してはある程度は正しかったと思います。もう1つの可能性は、実は目の前にあるのは雑多な現象で、何の体系もないのだけれど、そもそも人間の思考が体系的にしか理解できないようになっているので、何らかのシステムを抽出してしまわざるをえないということ。
我々が発生にそうしたシステムを見てしまうのには2つの可能性があると思います。1つは、実際に生きものが我々が感知したシステムどおりの序列で進化を遂げており、それを見てシステムと呼んでいるという可能性です。これは、分類学や系統学に関してはある程度は正しかったと思います。もう1つの可能性は、実は目の前にあるのは雑多な現象で、何の体系もないのだけれど、そもそも人間の思考が体系的にしか理解できないようになっているので、何らかのシステムを抽出してしまわざるをえないということ。
けれども、私たちにシステムの認識を教えたのは、自然だったのかもしれませんよね。原型という概念は人間が創出したものかもしれないけれど、自然界には、進化を経たことによる多様性があるわけで、それを見ているうちに自ずと人間は原型を作ってしまったのであれば、実は僕らは自然の被害者なのではないかと言ってみたいのです。原型を語ると観念論になりますが、原型を感知したことそのことの重要性もあるはずです。
まとめるなら、タイプライターの例で話したように、進化には外すことのできない保守的なパターンがあると思う。そのパターンの背景に、発生過程における細胞の出会いや組織間での相互作用、それによって発現する遺伝子のネットワークを見ていますが、それは何らかの形でゲノムの保守性とも関わっているでしょうね。だから、アノマロカリスゲノムの復元は挑戦する価値のある思考実験だと思うのです。
中村
私もまさに同じように考えていますが、倉谷さんに教えられるのは形の大切さですね。そこで遺伝子というより細胞の動きを重視する見方になる。そこ大事だと思うのです。
倉谷
細胞の三次元的な空間的配置、つまり細胞の出会いが鍵ですね。細胞数の多い動物ほど、どの細胞がどこで出会うかというのが決定的に重要になりますでしょう。
中村
細胞の出会いが、次々にカスケードを作っていくイメージですね。
倉谷
その通りです。センチュウならば、発生初期において数個の細胞から細胞系譜が割り出せます。
中村
ブレナー(註24)がセンチュウに注目したのは先を見る眼があったと思います。センチュウは細胞数も少なく細胞系譜がはっきりしているので、そういう形での発生の姿がよく見えてきましたものね。
倉谷
でも脊椎動物の発生における、咽頭胚の1個の細胞とセンチュウの1個の細胞を同じ重みで捉えることはできないこともわかってきた。やはり細胞と細胞の関わりによってダイナミックな形が生まれてくるのが脊椎動物であって、発生過程の1個の細胞だけを取り出して何の器官になるかを判断することは難しい。
中村
 確かにそう。道は一本ではない。その辺りが生きものに慣れていない人には非常に難しいところですね。たまたま細胞が出会ってある現象が起き、次はこうなってという連続ですね。
確かにそう。道は一本ではない。その辺りが生きものに慣れていない人には非常に難しいところですね。たまたま細胞が出会ってある現象が起き、次はこうなってという連続ですね。
倉谷
昔流の解剖学や形態学で、フナやカエルを通して勉強する方が、むしろ入っていきやすいでしょう。今の若い人は、その生きものを見たこともないのにいきなり細胞の絵を教科書でドーンと見せられてしまうのだから、訳がわからなくなってしまう。
中村
当たり前ですけど、一度はしっかり生きもの全体を見ておかなければいけませんね。
(註22) イデア【idea】
プラトン哲学の中心概念で、理性によってのみ認識される実在。感覚世界の個別の本質・原型。また価値判断の基準となる、永遠不変の価値。近世以降、観念また理念の意となる。
(註23) リンネ【Carl von Linn】
(1707-1778)
スウェーデンの博物学者。二名法を採用し、生物分類学の方法を確立。著書に『自然の体系』がある。
(註24) シドニー・ブレナー【Sydney Brenner】
(1927-)
イギリスの分子生物学者。1960年、フランスのF.ジャコブらと mRNAを発見。セントラルドグマの実体解明。線虫の分子生物学を始め、器官の発生と細胞死の遺伝学的研究によりノーベル生理医学賞受賞。
7.キャナリゼーションの集団発生学
倉谷
ヨーロッパにおける進化生物学の歴史を振り返ると、発生と進化がほぼ同義語として使われてきた自然哲学の経緯があり、その最後のあだ花がヘッケルの反復説だったとも言えるんですよ。今のエボデボ、進化発生学の1つの使命は、ヘッケルが間違った形で言ってしまったことを、理論的な機構論で読み替えること、そしてゲノムにつなげていくことです。
中村
まさにそうですね。ゲノムとつながってほしいんです。それが生命誌なので。
倉谷
発生学者は、ゲノムから発生を考える癖がありますが、僕はゲノムは到達地点だと思っています。ゲノムが成体の形をつくっているということは、成体の形を通した淘汰の中でゲノムが残ってきたということであり、つまり形態がゲノムに制約をかけているのです。例えば僕らの首、胸、腰は前後軸に沿って非常に整然と形態分化しているから、整然と並んだホメオボックス遺伝子の正しい発現が体作りの至上命令のように思える。ところが、同じ観点からヘビのようなちょっと変わった形の生きものを考えてみると・・・。
中村
胸と腹の順番なんてどうでもいいような気がする(笑)。
倉谷
ヘビのHOXコード、またはその元になっている遺伝子クラスターの構造的な保守性にかかる淘汰圧は、僕らのそれより緩いはずです。そうした例外的な形も含めてゲノムを考えるべきです。親の形を作るために必要なゲノムは、胚の形の取捨選択で選ばれているはずですね。そして、胚の淘汰を進化の原動力と仮定するなら、個体ではなく集団を扱わなければいけない。
中村
「集団発生学」ですか。
倉谷
それありですね。発生が本当に進化生物学の土俵に乗るためにはそれが必要ですし、その気になれば実現できるはずです。例えば、厳密に発生ステージを揃えた100個体の胚は、本当はどれだけ均一な集団なのか。ホールマウントin situ ハイブリダイゼーション(註25)や、あるいは何かの遺伝子のプロモーター領域につないだGFPを導入して遺伝子の発現領域とその強さにどのくらいのゆらぎがあるかを観察する。わずかな個体差がどのようなゆらぎ方で進むのか、変化の幅や勾配をきちんと計測できる実験系ができるはずです。
中村
研究館の小田さんが扱っているオオヒメグモは、それができるんです。母親にRMAを注入すると、それを発現した卵を200個くらい生んでくれる。
倉谷
いいなあ。ヤツメウナギも10万個くらい卵生みますが、遺伝子操作が中々できません。
中村
1910年代に発生生物学者、具体的には前館長の岡田節人先生の先生であるウォディントンが提唱したキャナリゼーション(註26)の考え方に取り組む時代になっていますよね。集団発生学はその具体化。
倉谷
ウォディントンの考えを察する研究者は今後増えていくでしょう。彼は、発生を坂道を転げ落ちる球になぞらえた(図3)。様々な遺伝子によって下からひっぱられた道は決して平らではなく、山あり谷ありですが、安定した場所で多少の外的擾乱やゲノムの変化が加わっても経路は乱れませんが、分水嶺に達した時は、わずかな揺らぎが大きく進路を変える。キャナリゼーションは、今の言葉を使えばゲノムが3%変わったからといって表現型が3%変わるわけではない。ゼロの時もあれば50%の時もあることを示したわけですね。
生体内で起こる小規模な変異、例えば高温のためにアミノ酸にごく小さな変異が起きた時などは、シャペロンのような熱ショックタンパク質(註27)のはたらきによって、タンパク質の形は保存されるわけで変異が中立化しますね。ところが外からの撹乱で、それまでに蓄積された中立的変異が全部表現型として噴出することもある。発生経路は安定化しているけれど、閾値を越えた撹乱が非線形の表現型の変化をもたらす可能性があるわけでしょう。
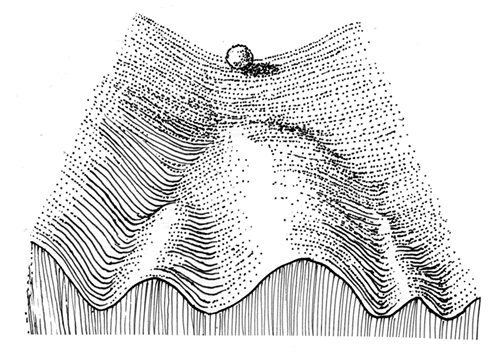
(図3)
後成的風景(epigenetic landscape)と呼ばれる発生を表す図。
中村
より具体的にはなんでしょう。
倉谷
昆虫の多型はその可能性が高いですね。ある種のシャクガの幼虫は、タンニンをわずかに多く摂取すると葉に擬態し、タンニンの量が足りないと花に擬態する。生きものが潜在的に複数の発生経路を持っているとすれば、多型こそが本来の姿と言ってもいいと思うんです。ゆらぎは安定化をもたらすと同時に進化の起爆剤になる。ここが面白いですよね。熱ショックタンパク質がはたらかなくなったショウジョウバエの変異体で、いろいろな表現型を起こす実験が成功した時は驚きました。変異がたくさん起きており、それをなんとか抑えて我々が生まれてこられるんだ、生きていけるんだということがわかったわけで。
中村
少し前の時代までは、進化は実験的な対象にならないとされて、第一線からひいた年寄りが考えることとされていましたが、発生とつながることで面白くなりましたね。「エボデボ」、進化発生学の時代ですね。
倉谷
当事者としては、これからどう盛り上がっていけるのか心配ですが。今やるべきことは、先ほど言われたように、胚が淘汰される対象であるということを見定めて、進化的変化を発生プログラムの変化として捉えて踏み込んでいくことだと思っています。
中村
それに、データの意味をきちんと考えていかなければいけませんね。昨年の対談でも、大規模な予算をとって行う実験を1年間お休みして、皆で紙と鉛筆を持ち寄ってデータをきちんと考える時間を持ってはどうでしょうと言ったのですが、これ本気なんです。
倉谷
私もそう思います。「スクール」は古代ギリシャ語では「暇」を意味する言葉でしたね。
中村
学校は暇、つまり余裕があるから行くところですよね。暇のない人は学校なんて行っていられない。
倉谷
 漁や野良仕事といった1日の仕事が終わって、「さあ今日は仕事が早く終わって暇だから、あの変なおやじのところに話を聞きに行こう」と、哲学者と呼ばれる人のもとに集ったのが大学ですね。この社会の中で大学くらいは、多少の暇と余裕をもって研究をしていてもよいと思うのです。
漁や野良仕事といった1日の仕事が終わって、「さあ今日は仕事が早く終わって暇だから、あの変なおやじのところに話を聞きに行こう」と、哲学者と呼ばれる人のもとに集ったのが大学ですね。この社会の中で大学くらいは、多少の暇と余裕をもって研究をしていてもよいと思うのです。
中村
説明責任が問われて、専門家として大事だと思うこと、先を見つめたことができませんね。
倉谷
研究者を目指す若い人は、他の学生と比べて自分にわがままであると同時に、自分にとっての理想にとても敏感です。彼らの最終目標である大学教授が雑務に忙殺されているというのはあんまりです。以前に一緒に岡山大学へ赴任した阿形さんともそういう話をしていて、彼は「学生の前では嫌そうな顔をしてはいけない。教授が嫌そうな顔をしていたら、学生は何のために努力をしようと思うんだ。」とよく言っていました。
中村
阿形さんは、どんなにつらくても楽しそうな顔をしている。それはすばらしいことだけれど、本当はもう少しゆっくり考えることができる状況にならなければいけないと思っています。
倉谷
アカデミズムをしっかり作り上げたのは、ヨーロッパ、特にイギリスやフランスですね。その国の学者が皆、偉大かというとそうでもなく、名前が残っている人は数えるほどです。
中村
それで良いわけでしょ。一人もいないのは困るけれど、ある割合ですばらしい仕事が出るためには、ゆとりが必要です。
倉谷
例えばニュートンやダーウィンには、彼らの知見に賛同して広めていく人も必要ですし、反対の立場から議論を重ねていく人も必要です。
中村
歴史に残る著名人はわずかでも、そこに関わる人がたくさんいたから、学問ができたのですよね。もちろん、よい仕事をしたいというのが研究者の願いですが、それはお金と効率で生まれるものではないということ。カメやヤツメウナギからの面白い結果を期待しています。

研究館実験室フロアーに掛けられたヘッケルの系統樹の前で。
(註25) ホールマウントin situ ハイブリダイゼーション【whole mount in situ hybridization:WISH】
調べたい遺伝子のmRNAに相補的なDNAで、生体内のどこにあるかを検出する。
(註26) キャナリゼーション【canalization】
C.H.ウォディントン(1905~75)によって提唱された発生過程の概念。発生過程は“運河化(canalized)”されており、外因(環境)や内因(遺伝的撹乱)の影響のもとでも、発生の進行はある決まった方向へと導かれるとする。
(註27) 熱ショックタンパク質【heat shock protein】
熱ストレスによって合成が誘導される一群のタンパク質で、HSPと略称する。熱によって変成したタンパク質を再び正しい立体構造に戻したり、逆に分解を促進する役割を持つ。遺伝子変異により構造が不安定となったタンパク質に結合し、正常な機能を発揮できる形になるように助けるはたらきを持つものもいる。
写真:大西成明
対談を終えて
倉谷滋
この対談は、息子が生まれたばかりの3月、そろそろ桜が咲く気配のする季節に行われた。「続く」という、生命進化の本質に関わるテーマは、同時に自分自身を構成するゲノムの半分が次世代に受け継がれる、この個人的イベントとも重ならずにはおれない。スイスの動物学者ポルトマンによれば、ヒトは子宮外において1年間胎児として過ごすというが、生まれた直後から発動を始めるべく新生児に準備されたさまざまな解剖生理機能を目の当たりにするにつけ、それは「巧妙」というには我々の想像力を遙かに超える。胚は外界から遮断されながらも、生まれ落ちてからでしか意味を持ちえない様々な適応的ロジックを自身の中にすでに抱え込んでいる。このような仕組みは、進化の単純な反復などから生まれようがない。むしろ発生プロセスそれ自体が、進化の産物なのである。

倉谷滋(くらたに しげる)
1958年大阪府生まれ。京都大学理学部卒業、同大学院修士・博士課程修了。理学博士。琉球大学医学部助手、熊本大学医学部助教授、岡山大学理学部教授を経て、2002年より理化学研究所発生・再生科学総合研究センター創造的研究推進プログラム形態進化チームリーダー、2005年より同グループディレクター。著書に『動物進化形態学』など。
.jpg)

.jpg)
.jpg)










