

今号テーマ
「変わる」から見えるワクワク感
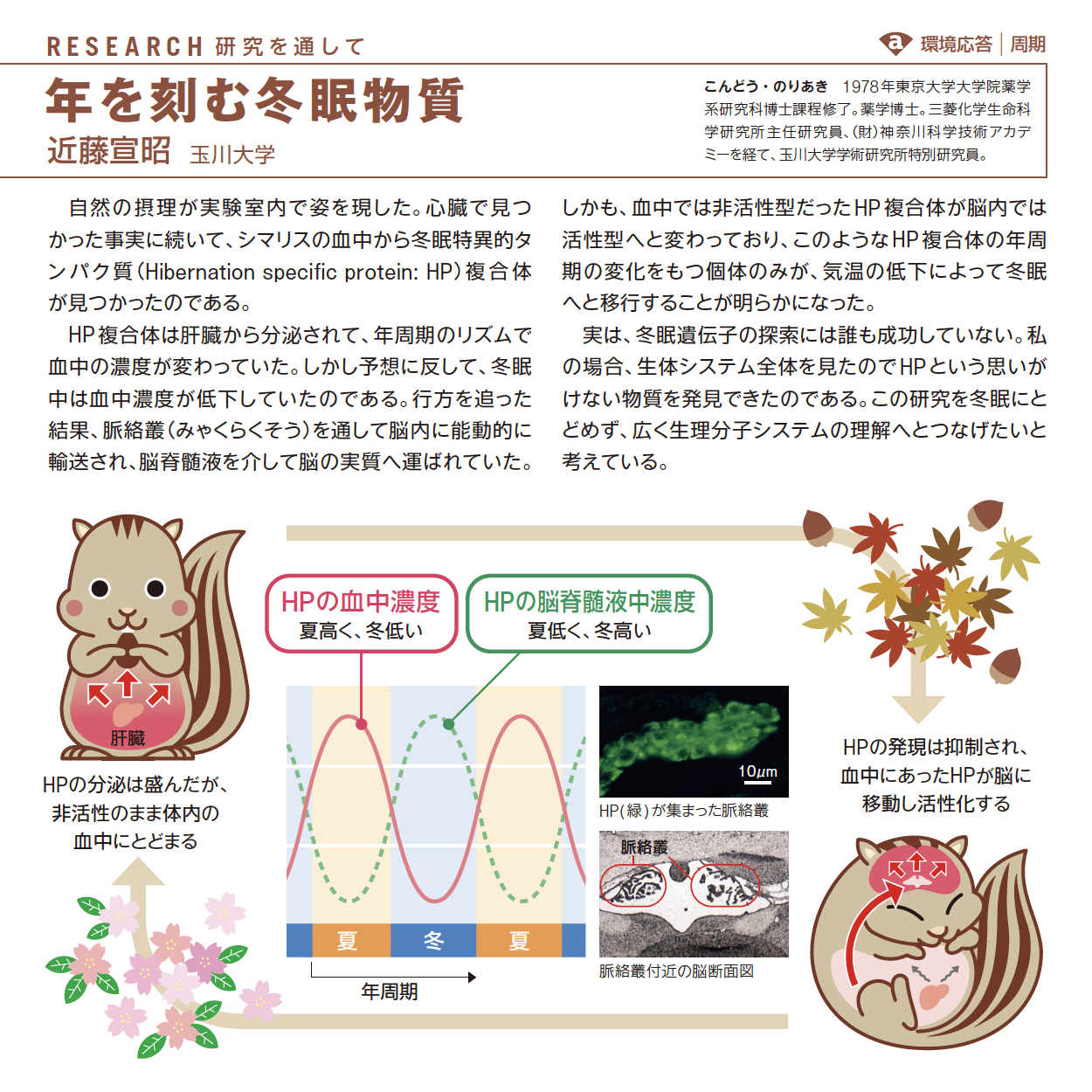
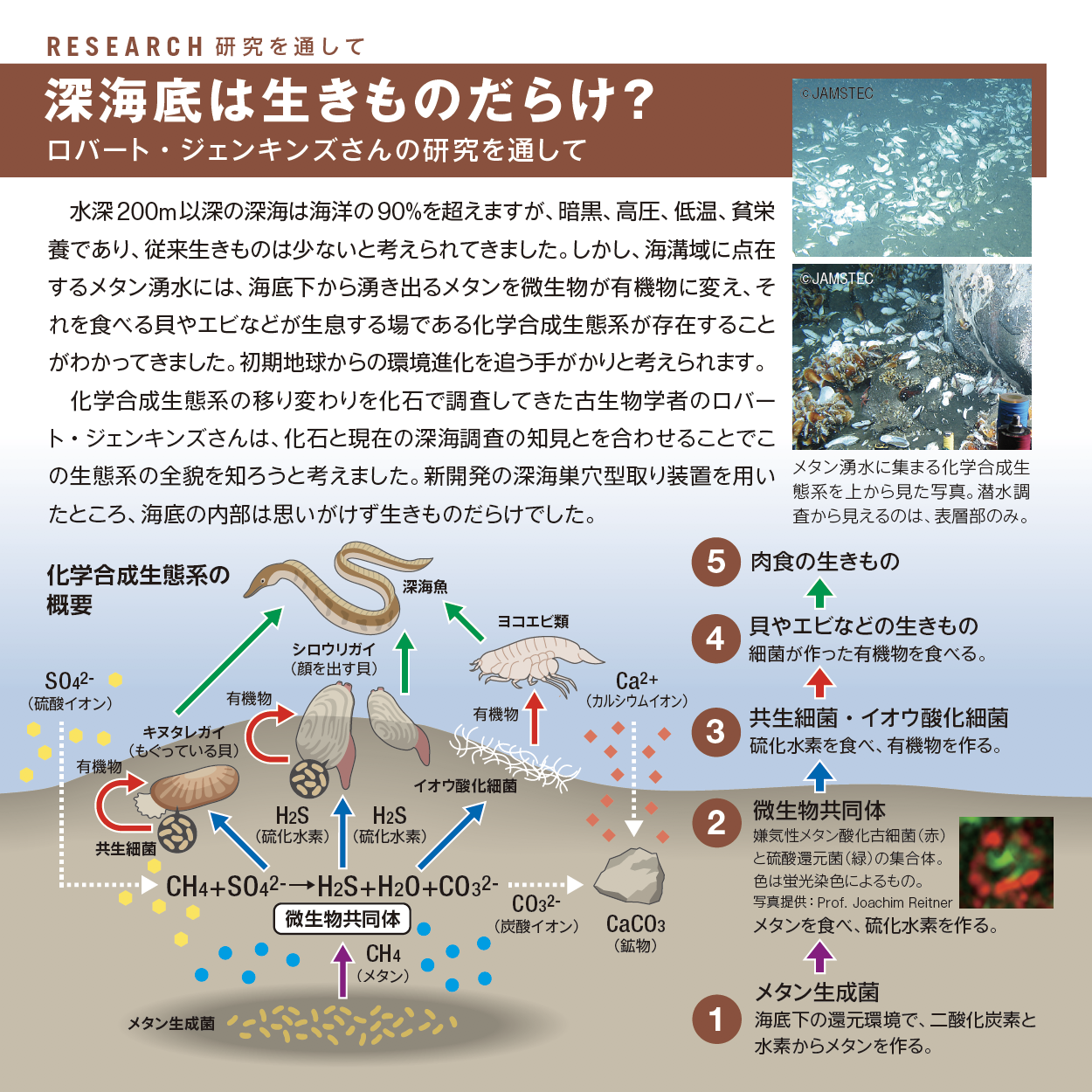
対談は、植物に囲まれている時の笑顔が魅力の長谷部さん。同じDNAを使いながら動物とは違う生存戦略をとり、地球生態系を支える植物の徹底研究のこれからが楽しみです。リサーチは、冬眠という身近な現象を追い続け、ついに冬眠物質を発見した近藤さん。研究者の執念を感じます。ロバート・ジェンキンズさんの海底巣穴型取りは過去を今そこに見せてくれてわくわくします。サイエンティスト・ライブラリーは分子生物学、免疫学という新しい学問との出会い、そこでの人との出会いを大切になさった谷口維紹さんです。(中村桂子)

TALK
植物の知恵に学ぶ
長谷部光泰基礎生物学研究所教授
中村桂子JT生命誌研究館館長
RESEARCH
研究を通して


CARD
記事のエッセンスが詰まったカード型の読みもの
カードサンプルを見るカードサンプルを見る















2012年年間テーマ
変わる
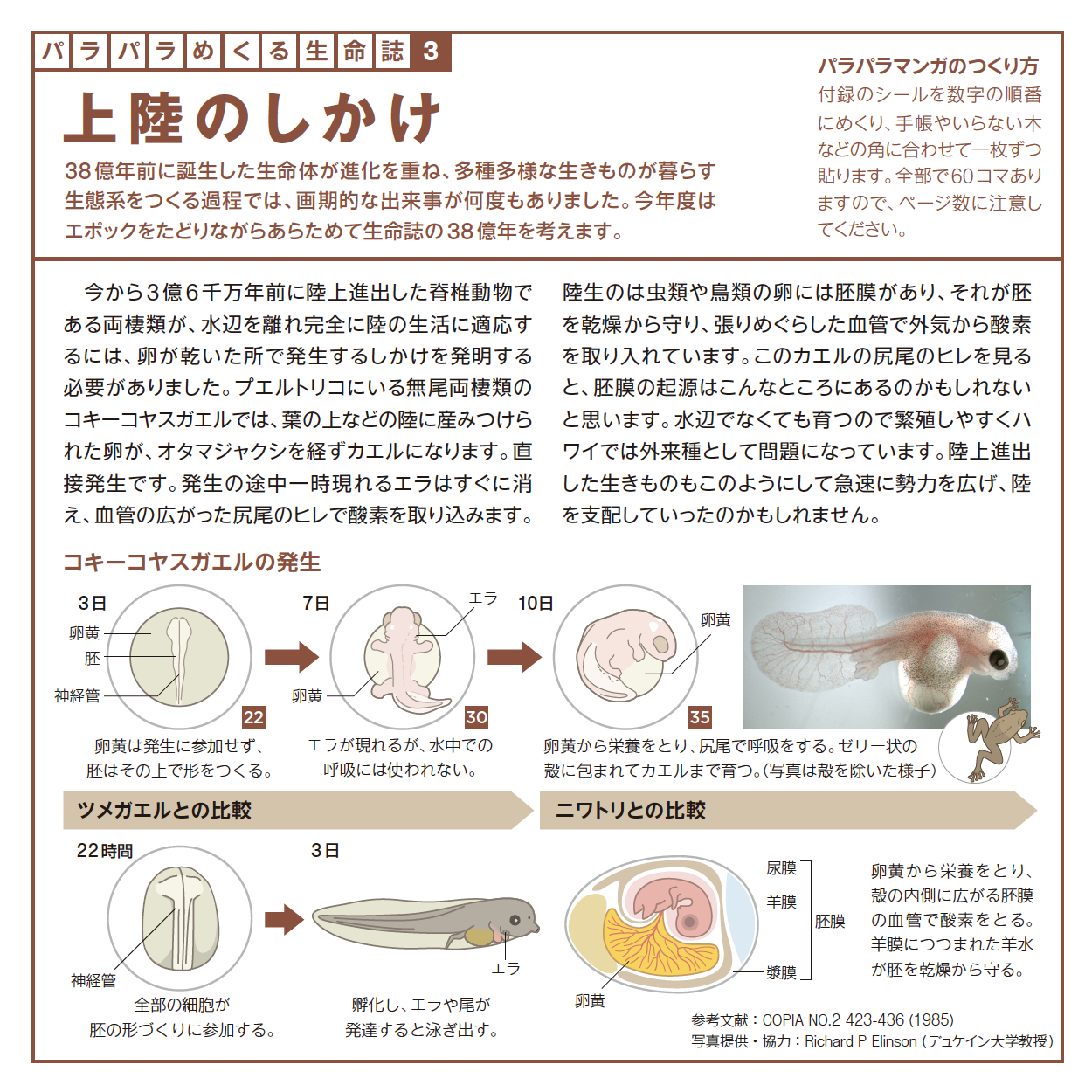
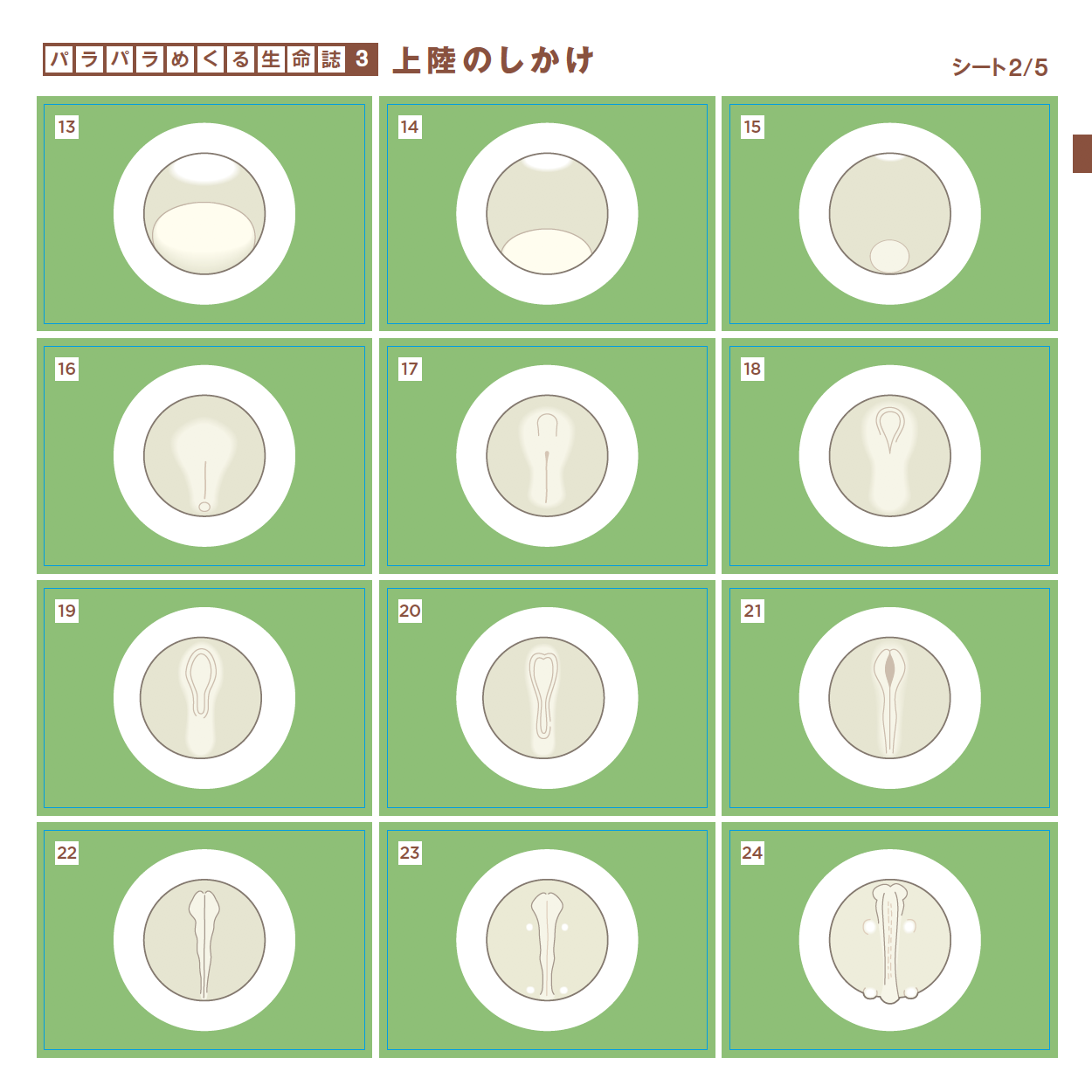
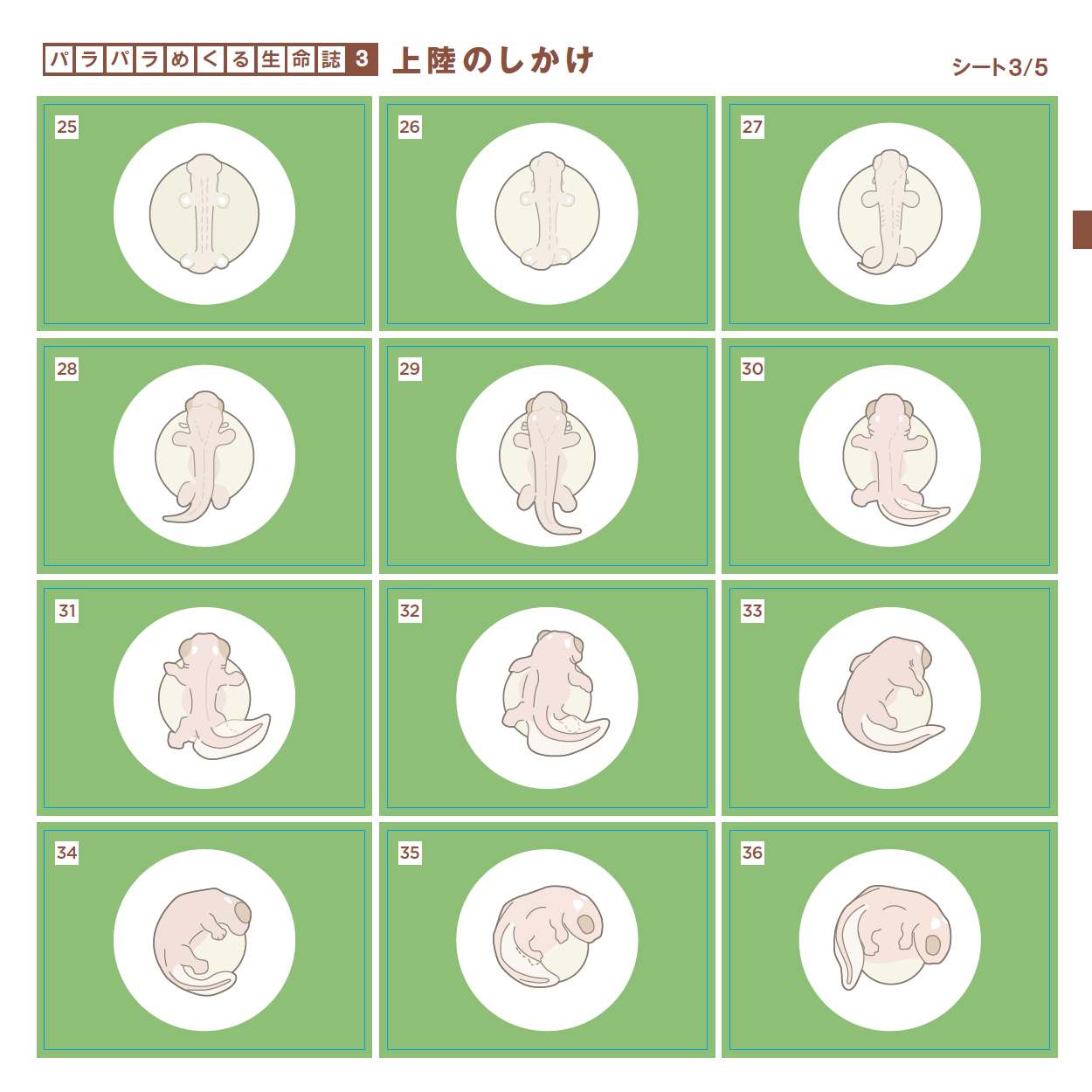
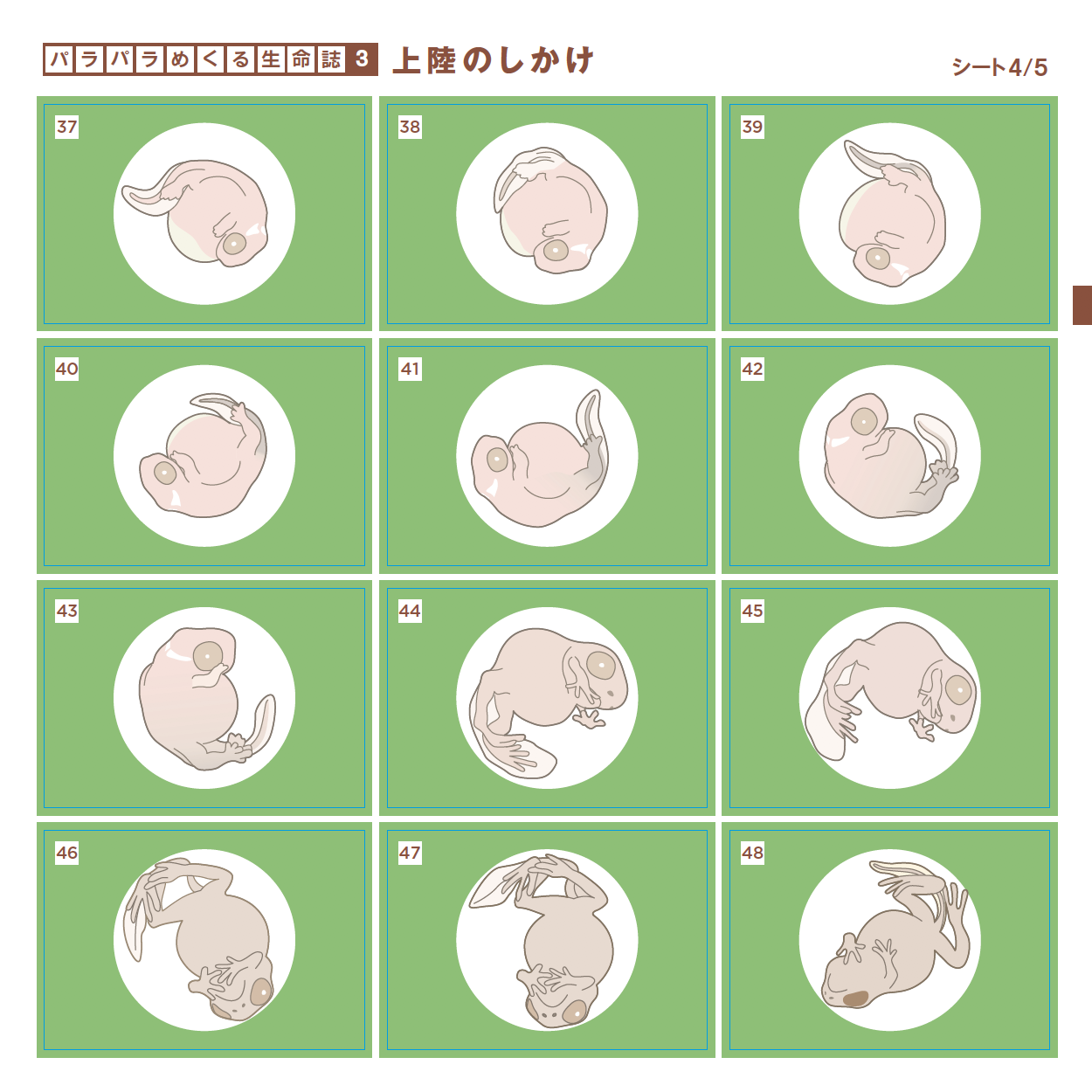
愛づる、語る、観る、関わる、生る、続く、めぐる、編む、遊ぶ・・・どれも「生きている」の大事な一面を表わす言葉ですが、今年は直球を投げ込みます。「変わる」です。つぼみだったバラがみごとな花を咲かせ、男の子がいつの間にかたくましい青年になる。それほどの変化でなくとも今日の私は昨日とは違っています。一方、そこには必ず変わらないものがあります。変わらないものを持ちながら変わる。38億年間それを続けてきたのが生きものです。社会も変わることを恐れてはいけませんが、底にある変わらないものも大切です。さまざまな現象に変と不変の微妙な組み合わせを見出しながら、「生きる」を考えます。


.jpg)

.jpg)
.jpg)










