

RESEARCH
形を生み出す相互作用
350年ほど前にロバート・フックが自作の顕微鏡を用いてコルク片を観察し、植物の組織が小部屋のような細胞でできていることを明らかにしました。植物細胞は動物細胞とは異なり、丈夫な細胞壁に囲まれていて、それが細胞の形を決めています。細胞壁の内側には細胞膜があり、その直下に微小管(表層微小管)が並んでいます。この微小管に沿ってセルロース微繊維が並び、細胞壁が作られるのです。微小管はすべての真核細胞に存在し、形を保ったり物質を運搬するなど重要な役割をしており、細胞内を移動していることが知られています。そのような微小管が細胞表層に並び、細胞壁を作り出すしくみはどのようなものか。東京大学の小田祥久さんは、道管を作る細胞の表層微小管に注目し、細胞壁の形作りを探りました。
1.植物の発生を支える細胞壁
生きものは細胞(cell)から作られている。350年ほど前にロバート・フックが自作の顕微鏡を用いてコルク片を観察し、植物の組織が小部屋(cell)のような構造の集まりであることを明らかにした。植物の組織を顕微鏡で観察すると、長方形あるいは多角形の細胞が整然と並び、確かに小部屋の並んだ建物のようだ。しかし、建物とは異なり、植物は成長し、子孫を残して枯れてゆく生きものである。静的なたたずまいとは対照的に、植物の内部では細胞が分裂と伸長をくり返し、動的に体を作り上げているのである。
植物細胞は動物細胞とは異なり、丈夫な細胞壁に囲まれており、それが細胞の形(図1)、さらには植物の形を決めている。細胞壁を完全に分解した細胞は球形になり、もはや植物の体を作れない。細胞壁で囲まれた植物細胞は移動しないので、植物の発生は個々の細胞の分裂と伸長の積み重ねで進む。細胞が移動・変形しながら発生する動物との違いである。分裂組織で増えた細胞は伸長して成長し、さまざまな形の細胞へと分化してゆく。
この過程は不可逆なので厳密にコントロールされていなければならない。植物を知るには細胞壁を知る必要があるわけだ。

(図1) さまざまな細胞の形を作りだす細胞壁
2.細胞壁は表層微小管に沿って作られる
細胞壁はセルロース微繊維と呼ばれる糖鎖を主成分とし、その間をさまざまな多糖類や糖タンパク質がつないでいる。細胞壁の内には細胞膜があり、その直下に微小管(表層微小管)が一方向に並んでいる(図2)。細胞膜に埋め込まれたセルロース合成酵素複合体が表層微小管に沿って動き、セルロース微繊維を合成するのである。したがってセルロース微繊維も一方向に並び、「たが」となって細胞の膨張を抑える。そこで、細胞はセルロース微繊維に対して垂直方向に伸びることになる。
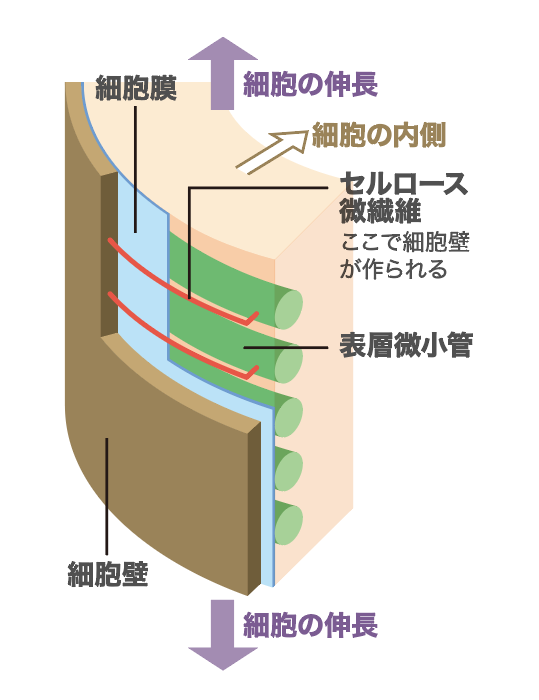
(図2) 細胞壁と表層微小管の関係
表層微小管の並びに沿ってセルロース微繊維が合成され、細胞壁ができる。
微小管は、すべての真核細胞に存在し、形を保ったり物質を運搬するなど重要な役割をしている。チューブリンというタンパク質が集合してできたチューブ状の集合体であり、チューブリンが結合したり離れたりすることで伸長と収縮を繰り返している。加えて、微小管同士が衝突したり、タンパク質で架橋して束になるなど、細胞内をダイナミックに移動していることもわかっている。しかし、細胞壁を作り出す表層微小管の並び方を制御するしくみは明らかにされていない。
3.道管の形成を観る
そこで、表層微小管の並び方の制御を知り、細胞壁ができるしくみを解き明かすために、私は根から吸い上げた水分を体全体に送る管である道管に着目した。道管は、通常の植物細胞がもつ一次細胞壁に加え、疏水的で丈夫な二次細胞壁を持つ。二次細胞壁の沈着後に、細胞内容物を消化し、細胞壁だけを残して中空構造になる(図3)。ただし、二次細胞壁は細胞の全面には沈着せず、壁孔と呼ばれる二次細胞壁の無い領域が残る。そこからまわりの細胞に水を行き渡らせ、道管としての役割を果たす。壁孔は規則正しく並び、らせん、網目、孔紋などの美しいパターンを形作る(図4)。実はこのようなパターンに並ぶのは表層微小管であり、それが壁孔の並びを誘導するのである。ここに着目し、表層微小管の並びを制御するしくみの解明に挑んだ。
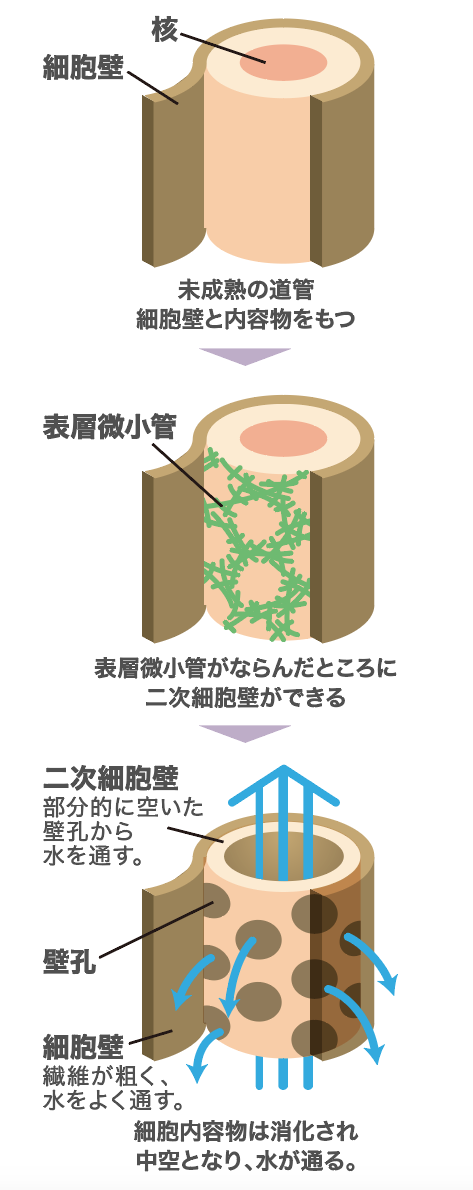
(図3) 道管における壁孔のでき方とそのさまざまなパターン
道管がもつ二次細胞壁は、細胞壁と同様に、表層微小管の並び方によってかたちが決まる。細胞内容物は細胞死によってなくなるため、道管は死細胞からできている。

(図4) 道管が作り出す壁孔のパターン
シロイヌナズナの木部の横断面。並んでいる管の一つ一つが道管。らせんや網目のような独自のパターンが見られる。
4.実験系の確立とMIDD1の発見
通常の植物細胞が道管へと分化する時の細胞内の表層微小管の振る舞いを明らかにしたいが、道管は植物組織の深いところで分化するので観察が難しい。そこで、モデル植物シロイヌナズナ(註1)の培養細胞を道管に分化させる方法を考え出した。まず、植物ホルモンによる分化誘導を試みたが、せいぜい30%程度の細胞しか道管を作らなかった。そのころ、道管への分化を誘導する遺伝子が発見されたので、さっそくそれを活用した。その結果、80%以上の細胞が網紋型の二次細胞壁を形成し、同調的に道管へと分化する系ができた。さらに遺伝子の効率的な導入法を確立して、細胞内のタンパク質を観察できるようにした。高性能の顕微鏡を用いて表層微小管の振る舞いを観察しているうちに、興味深いことに気がついた。表層微小管が壊れやすい場所があり、そこで次第に表層微小管が収縮し、壁孔ができていたのである。その時、表層微小管の分解を促進する未知のタンパク質があるに違いないと直感した。マイクロアレイ法(註2)を用いて、道管形成時にはたらいている遺伝子を調べ、そこから壁孔に局在するタンパク質を探索した。およそ1年にわたる探索の末、壁孔に局在し表層微小管に結合するタンパク質を見つけ、MIDD1 (Microtubule depletion domain1)と名付けた。予想通り、これが表層微小管の先端に作用して微小管を分解していた。
(註1) シロイヌナズナ
双子葉植物のモデル植物。高等植物の中で最初に全ゲノムが解読され、世界中の植物科学研究に利用されている。
(註2) マイクロアレイ法
多数のクローン(同一の遺伝的特性をもつ個体の集団)化されたDNA断片を高密度にプリントしたチップ(基盤)を用いて、生物個体の多数の遺伝子の発現を網羅的に解析する技術。
5.タンパク質(MIDD1)と表層微小管の相互作用による細胞壁形成
細胞内でMIDD1がはたらく場所はどのようにして決まるのだろうか。まず、MIDD1は低分子量GTPase(註3)の一つであるROP11を介して細胞膜につなぎ止められることがわかった。さらに、ROP11を活性化するROPGEF4および不活性化するROPGAP3が壁孔になる場所に集まり、ROPGEF4が存在する場所ではROP11が活性化され、MIDD1と結合していることもわかった。そこで、道管にならない植物細胞にROP11, ROPGEF4, ROPGAP3を同時に発現させたところ、確かにそれだけで局所的なROP11の活性化が起きた(図5)。そこにMIDD1を加えると、表層微小管が局所的に分解されて、二次細胞壁の道管にあるようなパターンができた。以上の結果から、ここで調べたタンパク質によって壁孔のパターンが作り出されていると考えてよさそうだ。
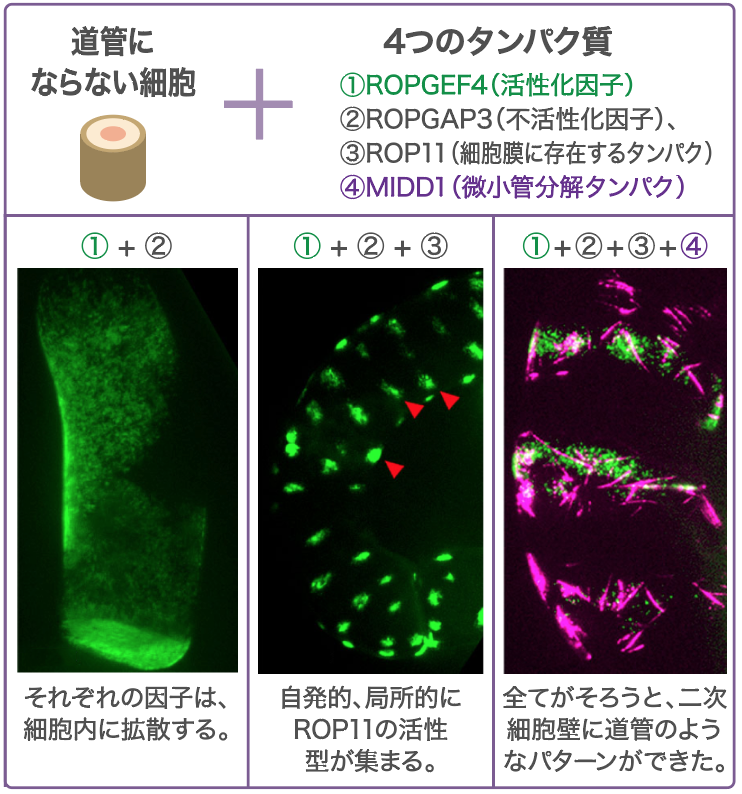
(図5) 壁孔のパターンを作り出すタンパク質
道管にならない細胞に、壁孔形成に関わるタンパク質を発現させ、その局在を見た。
これで、単純なパターンを持つ二次細胞壁を作り出すしくみは説明できた。しかし、網目やらせんなどの複雑なパターンを説明するにはこれでは不十分だ。一つのヒントが簡単な実験から得られた。微小管の構造を安定化する薬剤を加えたところ、壁孔が細長くなり、網目状に近いパターンの二次細胞壁ができたのである。表層微小管とROP11の位置関係を詳しく調べると、表層微小管が活性化したROP11の拡散を防ぐことで、壁孔のサイズを抑えていることがわかった(図6)。分子サイズの大きいMIDD1が、細胞膜につなぎ止められているROP11と結合して複合体を作り、移動を制限しているのである。
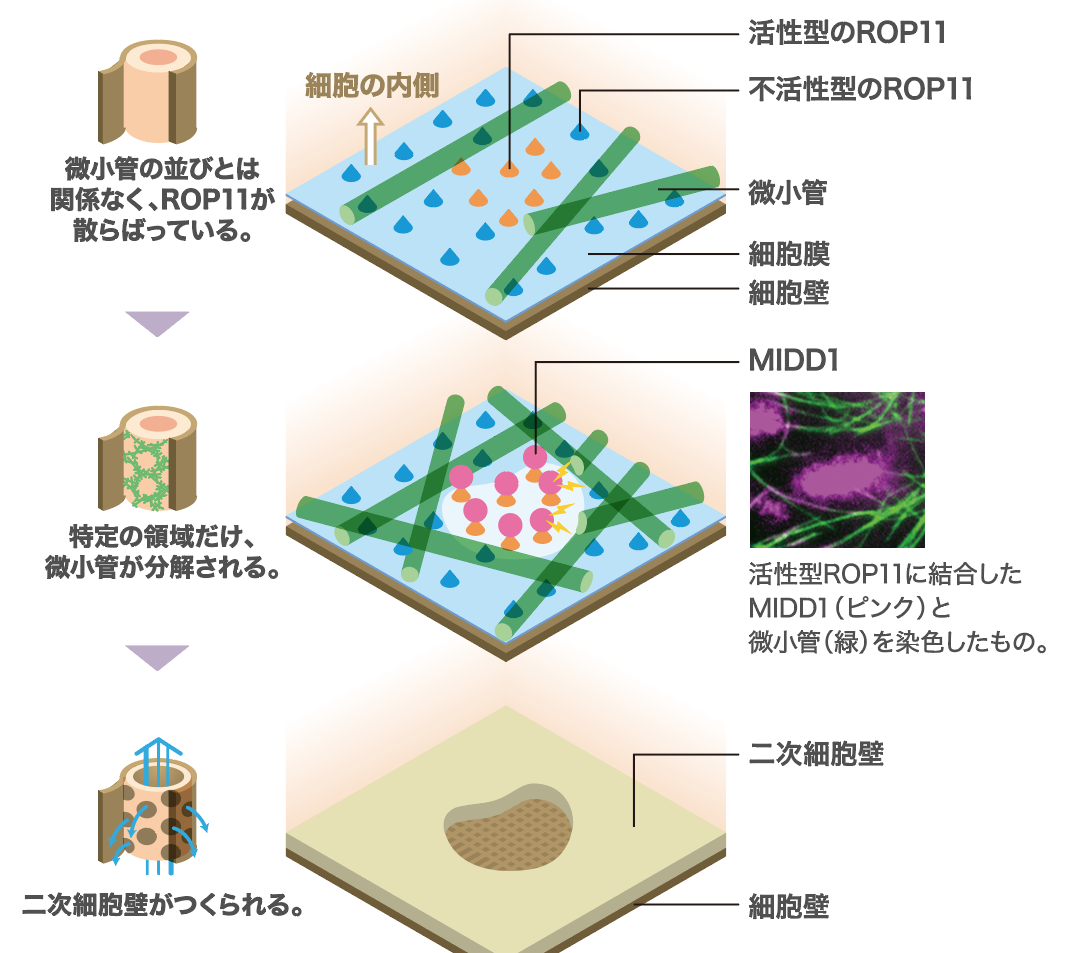
(図6) 道管の二次細胞壁を作り出す分子のしくみ
活性型ROP11に結合したタンパク質(MIDD1)と表層微小管の相互作用によって、壁孔は作られる。
(註3) 低分子量GTPase
GTPあるいはGDPと結合する分子量の小さいタンパク質の総称。GTPと結合すると活性型となり、さまざまな細胞応答を引き起こす。一方、GTPが分解されてGDPになると不活性型となる。こうした性質から分子スイッチともよばれている。
6.植物細胞の形作りの理解に向けて
道管をモデルとした研究により、局所的に活性化したROP11と表層微小管の排他的な相互作用が、細胞壁に壁孔のパターンをつくり出していることがわかった。局所的にROP11が活性化するしくみはまだ明らかではないが、チューリング・パターン(註4)と見てよいのではないかと考えている。興味深いことに、MIDD1とROP GTPase、そしてその活性化因子と不活性化因子は道管以外の細胞でも発現しており、類似したしくみが他の植物細胞の細胞壁でもはたらいている可能性がある。この研究を道管以外の細胞にまで広げてゆくことにより、植物細胞の形をつくり出す根本的なしくみを明らかにしたいと考えている。
(註4) チューリング・パターン
拡散速度の異なる活性化因子と不活性化因子が反応することにより、自発的に幾何学的なパターンが生み出される現象。動物の体表の模様をはじめ、さまざまな生命現象を説明することができるとして注目を集めている。数学者アラン・チューリングが提唱した。

小田 祥久(おだ よしひさ)
2007年東京大学大学院新領域創成科学研究科博士課程修了。博士(生命科学)。同年東京大学大学院理学系研究科研究員を経て、2011年より同大学助教と科学技術振興機構さきがけ研究者を兼任。
.jpg)

.jpg)
.jpg)










