

RESEARCH
発現調節配列の変化を探る
生きた化石といわれるナメクジウオは背骨を持たない脊索動物であり、私たち脊椎動物は、それと共通の祖先から進化してきたとされています。その過程では、全ての遺伝子が倍化する「全ゲノム重複」が2回起き、その時生まれた4個の遺伝子の機能が変わることで脊椎動物の複雑で多様なからだが作られるようになったと考えられてきました。
しかし近年、機能が変わっていない重複遺伝子がたくさんあることがわかってきました。違いを生み出すのは何か。山形大学の越智陽城さんは、遺伝子がいつ・どこではたらくかを決める発現調節配列の変化を追いました。
1.脊索動物から脊椎動物へ
生きた化石といわれるナメクジウオ(図1)は日本では瀬戸内海や三河湾に生息し、その一部は天然記念物に指定されている。一見サカナのようだが、体軸が脊索とよばれる柔軟な棒状の組織で支えられている脊索動物である。ヒトやカエルなど脊椎動物は脊索動物から進化してきたと考えられており、発生のごく初期には脊索で体が支えられていて、それがやがて脊椎に置き換わるのである。その進化の過程で、「全ゲノム重複」が起きていることが知られている。ここでからだ作りの遺伝子にどんな変化が起きたのだろう。

(図1) 明石沖で採取したナメクジウオ
体長は3~5センチ程度。生きている状態では、透明に近く内部が透けて見える。
2.全ゲノム重複と重複遺伝子のはたらき方の変化
脊椎動物の祖先種は、今から5億年以上も前のカンブリア紀にゲノムDNAが倍に増える「全ゲノム重複」を2回起こしたと考えられている。これによりすべての遺伝子が2度にわたって倍になったのだから、ゲノムに同じ遺伝子が4つづつ存在することになる。重複で余分に増えた遺伝子は、変異を起こして機能を失うか、時には新しい機能を獲得するかのいずれかの道を辿ると考えられてきた。このように、重複遺伝子の機能が変わることで、脊椎動物の複雑なからだ作りがなされるようになったとされてきたのである。ところが近年、ゲノムが解読された多くの動物種の重複遺伝子の比較や、ノックアウトマウス(註1)を用いて遺伝子の機能解析をしたところ、脊椎動物のゲノムには、機能が変わらず元と同じタンパク質を作っている重複遺伝子がたくさんあることがわかってきた。
そこで、可視化技術を用いて胚発生における重複遺伝子の発現を比べると、はたらく細胞や組織、はたらくタイミングが異なっていることが明らかになってきた。つまり、重複遺伝子の間で機能は変わっていないが、いつ・どこではたらくかが変わっているということである。遺伝子そのものではなく、遺伝子のはたらき方が変わることで進化が起きているのである。
遺伝子がいつ・どこではたらくかを決めるDNAを「発現調節配列」と呼び、そのはたらきは2種類ある。遺伝子を発現させるエンハンサーと、抑制するサイレンサーである。はたらき方の変化を見るには「発現調節配列」に注目する必要がある(図2)。
(図2) 全ゲノム重複と遺伝子の変化
全ゲノム重複によりタンパク質の設計図および、発現調節配列もすべて倍化する。ナメクジウオは全ゲノム重複を経験しておらず、祖先遺伝子に近い遺伝子をもつと考えられる。
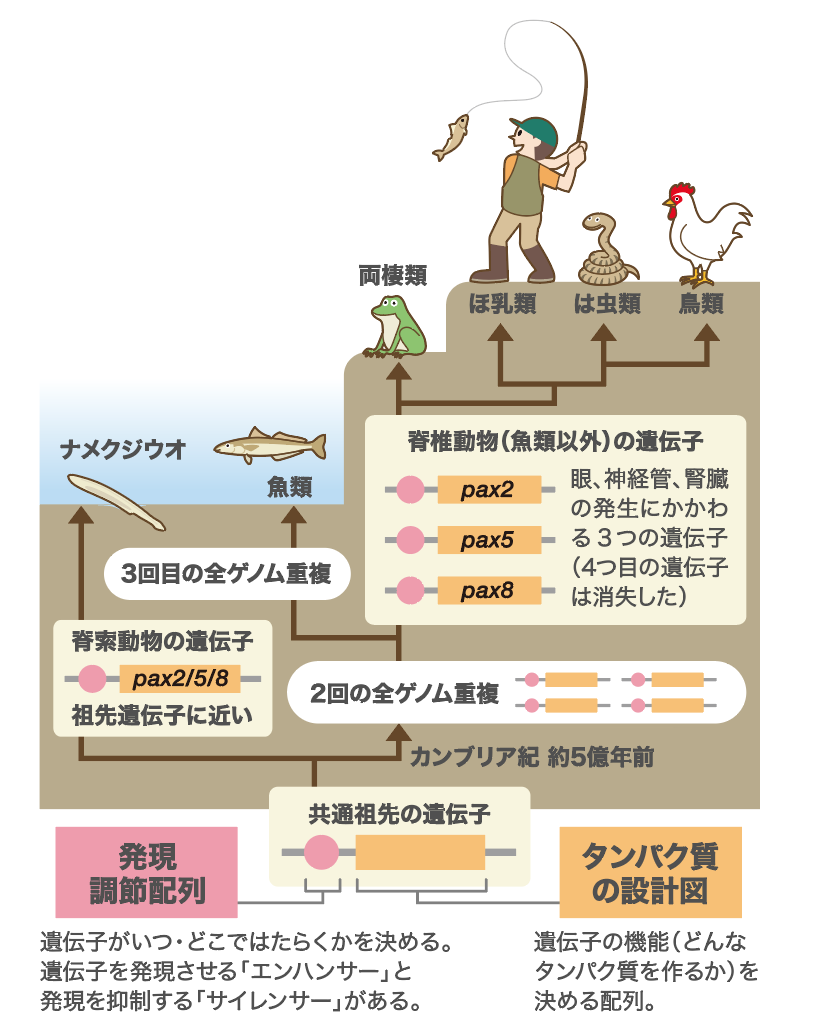
1999年に、オレゴン大学のAllan ForceとJohn Postlethwaitは、ゲノム重複でつくられた2つの遺伝子のはたらきを発現調節配列の変化で説明する、Duplication-Degeneration-Complementation (DDC:重複退行相補)モデルを提唱した(図3)。重複前の遺伝子が、組織Aで発現させるエンハンサー (緑色)と、組織Bで発現させるエンハンサー(青色)の2つをもっていたとする。そして、遺伝子の重複後に、一方は組織Bでのエンハンサーを失い組織Aでのみ発現し、もう一方は組織Aでのエンハンサーを失い組織Bでだけ発現するようになったとする。こうなれば、重複した遺伝子のいずれもがゲノムに残り、祖先遺伝子の発現様式が維持されることになる。近年このモデルを支持する研究成果が多く出ている。このモデルは配列の変化で既存のエンハンサーが失われる例だが、発現の多様化には新たな機能を獲得する例もあるのではないだろうか。そう考えて、私たちはこの問題の解明に挑んだ。
(図3) 配列の変化で既存のエンハンサーが失われる例
Duplication‐Degeneration‐Complementation(DDC:重複退行相補)モデル。祖先遺伝子が2つの組織(AとB)で発現させるエンハンサーをもっていると、ゲノム重複後、それぞれの遺伝子座でいずれかのエンハンサーを失っても、両遺伝子が発現するので祖先遺伝子と同じはたらきをもつことができる。
3.エンハンサーのはたらきを見る
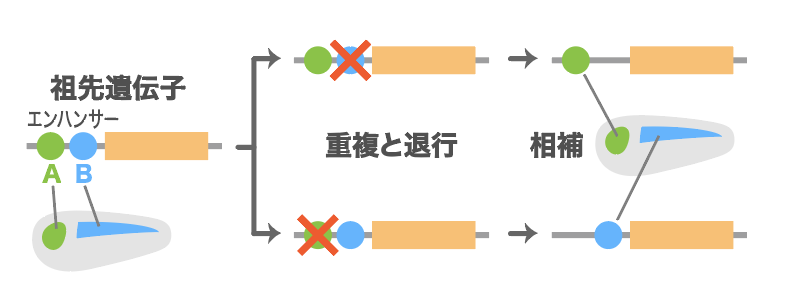
私たちは脊椎動物ではたらく2つの遺伝子Pax8とPax2に注目した。これらは全ゲノム重複によって生まれた遺伝子で、Pax8は主に腎臓で発現し、Pax2は眼、中脳後脳境界(神経管)、腎臓で発現する。ナメクジウオは、Pax8とPax2の共通の祖先遺伝子に近いPax2/5/8を持ち、それは脊椎動物のPax2と同じように、原始的な眼(眼点)と神経管 (神経索)、腎臓 (腎管)で発現する(図4)。
(図4) Pax8、Pax2、Pax2/5/8の発現場所
Pax8だけが腎臓でのみ発現する。
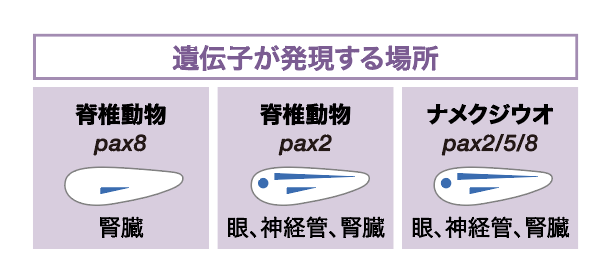
そこで、Pax8とPax2それぞれのエンハンサーのはたらく組織を調べた。まず、蛍光タンパク質をつくるGFP遺伝子にPax8とPax2のエンハンサーをつないだ(実際にはエンハンサーとGFP遺伝子の間に基本プロモーターと呼ばれる配列を入れなければならないがここの話ではこれは忘れてよい)。これをツメガエルの胚に入れ、どこの臓器・組織が光るか、つまりエンハンサーがどこではたらくかを見たのである(図5)。すると、いずれの場合もGFP遺伝子は眼と神経管、腎臓で発現した。これまでに知られているエンハンサーは、眼での発現、腎臓での発現というように臓器毎に役割が振り分けられていることが多いのだが、Pax8とPax2のエンハンサーは眼と神経管と腎臓という、複数の組織で遺伝子を発現させる多面的な機能をもつことがわかった。
Pax8とPax2のエンハンサーのはたらきがよく似ているので、配列を比べてみたところ、それも非常によく似ていることがわかった。つまり、Pax8とPax2のエンハンサーは、祖先遺伝子がもっていたものが、全ゲノム重複の後も残っていたと考えられる。そこで、祖先遺伝子に近いナメクジウオのPax2/5/8のエンハンサーを調べたところ、やはり眼と神経管、腎臓ではたらいた。Pax8とPax2はどちらも祖先遺伝子と同じ機能のエンハンサーをもち続けていたのである。では、なぜPax8は腎臓でしか発現しないのか。ここに新しい変化を知る可能性がある。
(図5) エンハンサーのはたらき
GEPが発現する場所が青紫に染色されている。この結果から、Pax8、Pax2、Pax2/5/8のもつエンハンサーのはたらきは同じであることがわかった。
4.サイレンサーの探索
ここで私たちは、Pax8が遺伝子の発現を抑えるサイレンサーを獲得したのではないかと予想した。サイレンサーは発現の始まりを合図するプロモーター領域に存在することがある。そこで、蛍光タンパク質の発現を調べるときに用いた、基本プロモーターに変えてPax8自身のプロモーターをここに入れた(図6)。すると、蛍光は本来のPax8のように腎臓だけで見えた。さらに、ここにPax2のエンハンサーを入れても、Pax8と同じように腎臓だけで発現した。予想通りPax8にはサイレンサーが存在していたのである。これらの結果から、Pax8とPax2はいずれも同じエンハンサーを祖先遺伝子から引き継いでいるが、Pax8は眼と神経管でエンハンサーのはたらきを打ち消すサイレンサーを獲得したことがわかった(図7)。
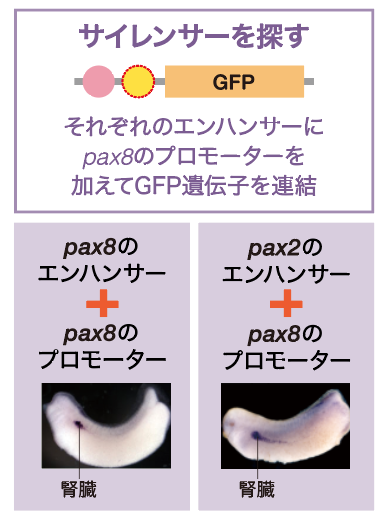
(図6) サイレンサーの探索
Pax8のプロモーター領域に、サイレンサーが存在することがわかった。
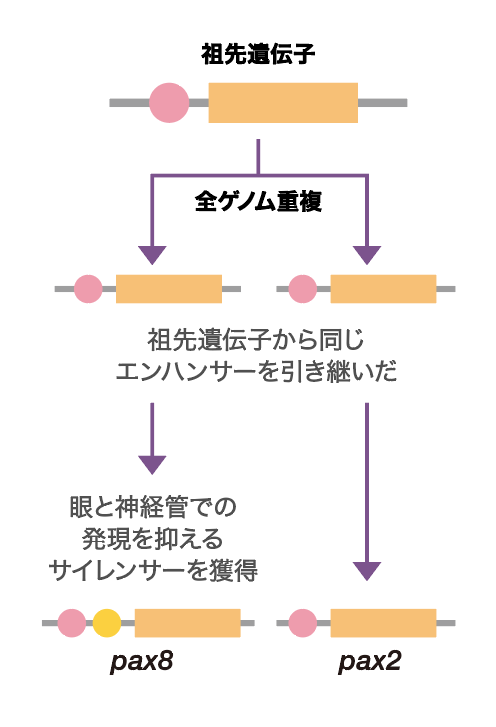
(図7) 発現調節配列の変化
5.サイレンサーの獲得が示す重複遺伝子の発現の多様化
これまで、ゲノム重複後の遺伝子発現の多様化の研究は、発現を促すエンハンサーに注目して行われてきたが、それだけでは現実の重複遺伝子の発現の多様さを説明できない。これに対して私たちは、サイレンサーの獲得が重複遺伝子の発現の多様化を生み出すという新たな切り口を示した。つまり、5億年以上も昔の祖先動物で既に完成していたエンハンサーが、遺伝子のコピー数の増加後、増えた遺伝子が余計な場所でははたらかないようにサイレンサーを獲得するという進化過程を明らかにしたのである。
謝辞
奈良先端科学技術大学院大学 荻野 肇 研究グループリーダー長、ならびにナメクジウオの採集にご協力いただいた兵庫県立農林水産技術総合センター 主席研究員 谷田圭亮先生に心から感謝致します。

越智陽城(おち はるき)
2003年奈良先端科学技術大学院大学(NAIST)バイオサイエンス研究科博士課程修了。博士(バイオサイエンス)。オレゴン大学研究員、NAIST助教を経て、山形大学医学部 教育研究支援センター助教。
.jpg)

.jpg)
.jpg)










