

RESEARCH
胎盤の多様化と古代ウイルス
エンベロープタンパク質が結ぶ
母と子の絆
哺乳類の胎盤は、母子間で栄養やガスなどの交換を行い、胎児の成長を助けています。哺乳類の器官の形は種間でよく似ていますが、胎盤の形だけは種ごとに大きく異なります。この多様性の原因は今も謎です。宮沢孝幸さんが注目するウシ科には、胎盤で母と子の細胞が融合するという特徴をもつ種がいます。ウシなどが属するウシ亜科とヤギなどが属するヤギ亜科です。この特徴の鍵を、ゲノムにある大昔に感染した古代ウイルスの痕跡が握っていました。胎盤の多様性は、有胎盤哺乳類の祖先と古代ウイルスの関わりで語れるのではないかと考えての研究が始まりました。
1.胎盤の役割と多様性
胎盤は有胎盤哺乳類の妊娠期に形成される器官で、妊娠中に子宮内で胎児を支えたり、母子間での栄養やガスの交換を行ったりすることで、胎児の成長を助ける役割を担っている。有胎盤哺乳類の他の器官や組織の形はよく似ているが、胎盤の形は動物種ごとに大きく異なる。例えば、ヒトやマウスの胎盤は円盤状(盤状胎盤)だが、イヌやネコは帯状胎盤、ウマやブタは散在性胎盤、ウシやヒツジは叢毛性多胎盤である(図1)。同じ形の胎盤でも、その組織学的な構造は動物種によって異なっている。このような胎盤の多様性の原因は、現在も大きな謎のままである。
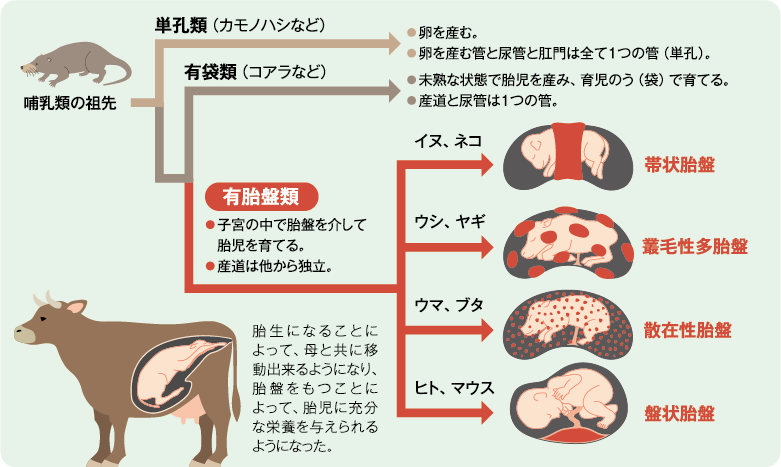
(図1) 哺乳類の生殖器官と胎盤の多様性
哺乳類は、胎生になることによって、母と共に移動できるようになり、胎盤をもつことによって、胎児に充分な栄養を与えられるようになった。胎盤の多様化の原因は、まだ完全には明らかになっていない。
2.ウシ科動物の胎盤の特徴
私たちは、特にウシやヒツジを含むウシ科動物の胎盤に着目して研究を行っている。ウシ科動物はウシ亜科、ヤギ亜科、インパラ亜科など7つの亜科に分かれるが、どの亜科の胎盤も多数の胎盤節からなる叢毛性多胎盤である。組織学的には胎盤は胎児側と母体側に分かれ、胎児側の胎盤には栄養膜単核細胞(MTC: Mononucleate trophoblast cell)と栄養膜二核細胞(BNC: Binucleate cell)という特徴的な細胞がある(図2)。そのうちBNCが、母体側の胎盤の表面にある細胞(子宮内膜細胞)と融合することで、母体へ侵食してホルモンを母体へ輸送し、妊娠を維持する役割を果たしている。
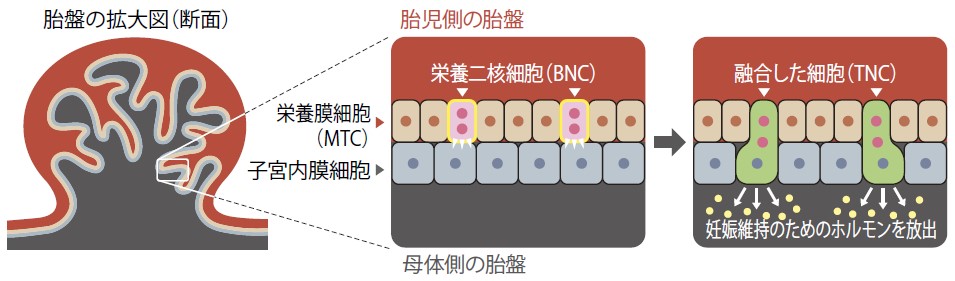
(図2) ウシ亜科動物の胎盤の断面図と細胞融合
細胞融合の形式は動物種によって異なり、ウシやスイギュウなどが属するウシ亜科とヤギやヒツジなどが属するヤギ亜科のみに、胎児側と母体側の細胞が融合する極めて特徴的な現象が見られる。ウシ亜科動物では、BNCと子宮内膜細胞が1対1で融合して三核細胞(TNC: Trinucleate cell)を形成し、ヤギ亜科動物では複数のBNCが1つの子宮内膜細胞と融合して多核細胞を形成する。ウシ亜科動物の胎盤におけるこのような細胞融合の特徴は、いったい何に起因しているのだろう。これまで研究方法がなく解かれていないこの問いをゲノムから攻めることを考えた。
3.ゲノムに埋め込まれた古代ウイルス
ヒトゲノムを解読したところ、レトロトランスポゾンと呼ばれる領域が約50%を占めることがわかっている。その中には逆転写酵素をもつレトロウイルス由来のLTR(Long terminal repeat)型レトロトランスポゾンと呼ばれる領域があり、それがゲノム全体の約10%を占めている(図3)。LTR型レトロトランスポゾンの中には、祖先の生殖細胞に感染したレトロウイルスがゲノムの一部となった古代ウイルスの名残りがある。これを内在性レトロウイルス(ERV)と呼ぶ。ERVのうち、ヒトではウイルスのエンベロープタンパク質に由来するシンシチン1,2が胎盤で特異的に発現し、そのはたらきによって胎児の栄養膜細胞同士が融合することが知られている。
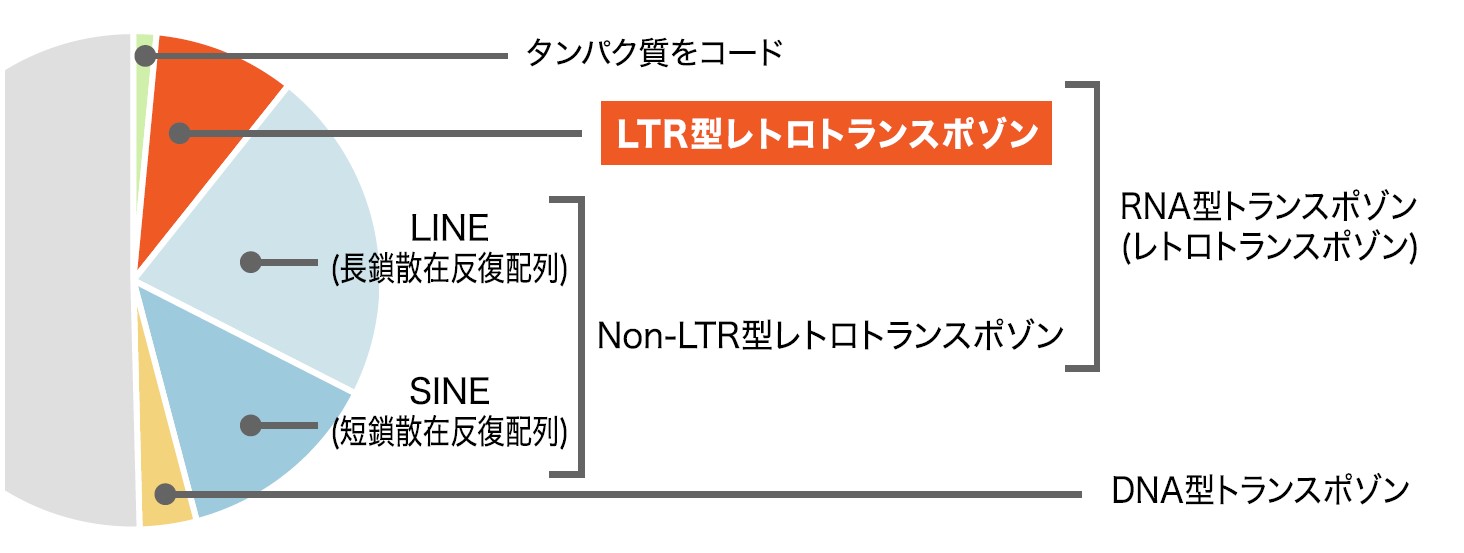
(図3) ヒトゲノムの構成要素
私たちがウシ胎盤を調べたところ、ウシ内在性レトロウイルスK1(BERV-K1: Bovine endogenous retrovirus K1)のエンベロープ遺伝子(env: Envelope gene)が発現していることが明らかになった。エンベロープタンパク質は本来、ウイルスが宿主の細胞に侵入するときに、ウイルスの外膜と宿主の細胞膜を融合させるはたらきをするものである。そこで、ウシ胎盤での細胞融合を特徴付けているのは、BERV-K1 のエンベロープタンパク質(Env: Envelope protein)と考え、ウシ胎盤で特異的に見られるこの配列と、TNC形成の関わりを調べた。
4.ウシ内在性レトロウイルスK1の役割
まず、BERV-K1 Envに結合する抗体を用いて、ウシ胎盤を免疫組織化学的に染色したところ、BERV-K1 Envは胎児側胎盤のBNCで特異的に発現していた(図4左)。次に、BERV-K1 Envの細胞融合能を確かめるために、BERV-K1 Envを発現させた細胞と母ウシ由来の子宮内膜細胞を一緒に培養したところ、高い頻度で細胞の融合が観察できた(図4右)。実際にウシ胎盤でも、BERV-K1 Envを発現したBNCが子宮内膜細胞と融合することを示唆する結果である。
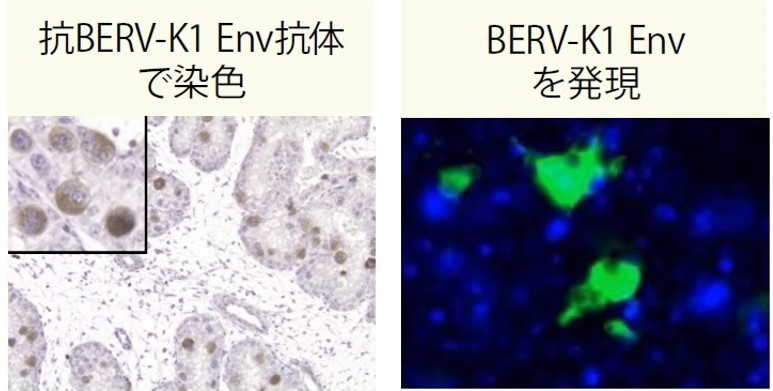
(図4)ウシ胎盤でのBERV-K1 Envの発現(左)とBERV-K1 Envによる細胞融合(右)
(左)BERV-K1のエンベロープタンパク質(Env)に結合する抗体を用いて、ウシ胎盤でのFematrin-1の局在を観察した。濃い部分がFematrin-1の在る部分。
(右)緑色に光る蛍光タンパク質(Kusabira-Green)を目印として、BERV-K1 Envが発現した細胞と子宮内膜細胞の融合を観察した。
他のウシ科動物でのBERV-K1 env遺伝子の有無を調べたところ、バリギュウ、アジアスイギュウ、シタツンガなどウシ亜科動物にしかなく、近縁なヒツジやヤギなどのヤギ亜科動物には無いことが確認された。これは、ウシ亜科動物とヤギ亜科動物が分かれたおよそ2000万年前に、ウシ亜科動物の祖先動物にBERV-K1が感染したことを示唆している(図5)。
ウシ亜科動物すべてのBERV-K1 env遺伝子は、純化選択によって保存されているので、BERV-K1 はウシ亜科動物の祖先動物のゲノムに入り込んだ後、比較的早い時期に、TNC形成能を獲得したと考えてよいだろう。ERVにおける系統樹の解析から、BERV-K1 Envはヒトの胎盤で発現しているシンシチンとは由来が異なり、ウシ科に特有のものだったので、私たちはBERV-K1 Envを母子間細胞融合誘導因子1(Fematrin-1: Fetomaternal trinucleate cells inducer 1)と命名した。
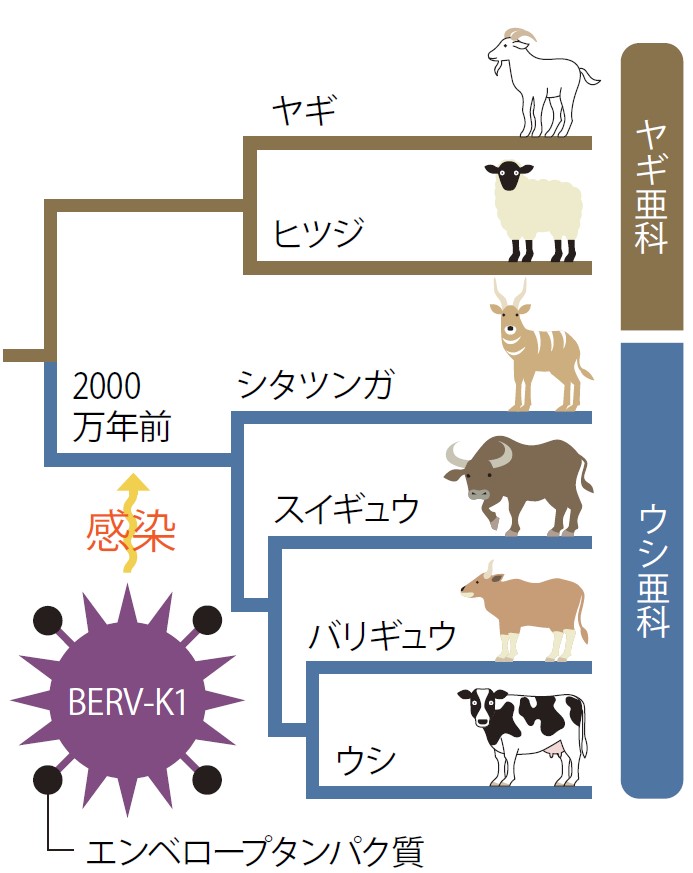
(図5)BERV-K1の感染時期(推定)とヤギ/ウシ亜科の系統樹
5.胎盤形成と古代ウイルス
こうして、ウシ亜科動物におけるTNCの形成機序が明らかになった。私たちは、他のウシ科動物にもFematrin-1とは異なる内在性レトロウイルスのEnvが存在し、同様に細胞融合の役割を担っていると予想している。そして、それぞれのEnvの性質が異なるため、動物種間での細胞融合の形式に違いが生じているのではないかと考えている。
ところで、なぜ様々な内在性レトロウイルスが胎盤形成に関わっているのだろうか。私たちは以下のような仮説を立てている。哺乳類が胎盤という新しい器官を生み出した当時、様々なレトロウイルスが哺乳類に蔓延し、ゲノムに組み込まれた。胎盤では、母体から胎児への攻撃を抑制するために、免疫抑制能をもつEnvが使われるようになったのであり、その中にはシンチシンやFematrin-1のような融合活性をもつものもあった。そして長い時間をかけた胎盤の進化の中で、様々なレトロウイルスのEnvが試され、生殖に有利なものが選択されて、現在に至ったのではないか。この仮説の検証が今後の仕事である。
私たちは現在、胎盤の多様化とERVの内在化の過程を明らかにするために、有袋類の仲間であるコアラのERVを含めた研究を進めている。

宮沢孝幸(みやざわ・たかゆき)
1990年東京大学大学院農学生命科学研究科博士課程獣医学専攻修了。博士(獣医学)。
英国ロンドン大学名誉研究員、日本科学技術振興事業団さきがけ21研究員、帯広畜産大学畜産学部獣医学科助教授などを経て、2009年より京都大学ウイルス研究所細胞生物学研究部門(信号伝達学研究分野)准教授。2013年より同研究所進化ウイルス研究領域准教授を兼任。
写真:胎盤の多様化を解くための鍵になるかもしれないコアラとのツーショット。2013年に、王子動物園で。
.jpg)

.jpg)
.jpg)










