

RESEARCH
巨大ウイルスから見える新たな生物界の姿
これまで、ウイルスは生きものではないとされてきた。ところが2003年に、ミミウイルスと呼ばれる巨大ウイルスが同定され、ウイルスの概念が変わった。巨大ウイルスがどのように進化し、生物界でどのような役割を担っているのか。そこから新しい生物界のありようを解明したいと考えている。
1.これまでのウイルス観
地球上の生きものは、大きく真正細菌、古細菌、真核生物という3つのグループ(ドメイン)に分類される。ウイルスはどの生きものにも感染し、真正細菌に感染するものをバクテリオファージと呼ぶ。中心にはDNA あるいはRNAのどちらか一方の核酸があり、それがカプシドと呼ばれる殻に包まれ、ウイルスによっては、さらに外側をエンベロープと呼ばれる膜が覆っている。
生きものに感染したウイルスは、細胞内でウイルス核酸の複製とウイルスタンパク質の合成を行い、それらが集まって新たなウイルス粒子を形成し、細胞から出て行く。動植物から細菌にいたる全ての細胞が二分裂で増えるのに対し、ウイルスは組み立て方式によって細胞内で増殖するのである。この時に使われるエネルギーやタンパク質は、感染した宿主細胞の機能を利用してつくり出す。そのため、ウイルスは生きものとは見なされてこなかったのだが、近年の研究によって新しい考え方が必要になってきた。
古くは「ろ過性病原体」と呼ばれていたように、多くのウイルスは100nm程度の大きさしかなく、光学顕微鏡で観察できない。ほとんどのウイルスのゲノムサイズは、真正細菌(約0.15—12Mbp)と古細菌(約0.5—6Mbp)より小さく、例えば大きいとされている単純ヘルペスウイルスのゲノムサイズでも約0.15Mbp(遺伝子数は約80個)である。ところが、2003年にミミウイルスと呼ばれる巨大ウイルスが同定されることによって、これまでのウイルス観が変わった(図1)。
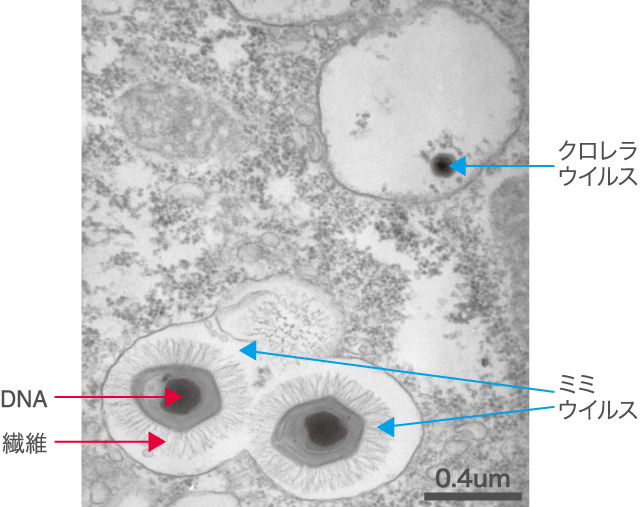
(図1)
アカントアメーバに取り込まれ、食胞の中に収まった2つのミミウイルスと、別の食胞の中に収まったクロレラウイルス。ミミウイルスの中心にはDNAの固まりが、周りには糖タンパク質と考えられている固く長い繊維が観察できる。写真:Chantal Abergel博士/CNRS-AMU提供。
2.常識を覆す巨大なミミウイルス
ミミウイルスは、1992年にイングランド北部の病院内で見つかった。病院ではアメーバを使って肺炎の原因になるリジオネラなどの病原性細菌探索という作業が行われており、その際に空調機冷却水サンプルからアカントアメーバにより増幅・単離されたのである。光学顕微鏡で観察出来るほど大きく、グラム染色されることから初めはグラム陽性菌の一種と考えられ、発見された町名にちなんで当初はブラッドフォード球菌と名付けられた。しかし、そこにはタンパク質合成に関わるrRNA遺伝子がなく、細菌としての特徴付けは難航した。
それから10年ほど経った2003年、フランスの細胞内細菌の専門家・ディディエ・ラウルトのグループにサンプルが渡り、電子顕微鏡での詳細な観察から、それが巨大ウイルスと結論付けられた。本体の直径は約0.4μmでエンベロープはなく、正二十面体のカプシドに包まれていることが分かった。カプシドには糖タンパク質と考えられる長い繊維が付いており、それを加えると直径約0.75μmもあった。それまで巨大ウイルスとされていた天然痘ウイルスやクロレラウイルスの直径が約0.2μm、小型の真正細菌・マイコプラズマの直径が約0.3μmであるから驚きの大きさである(図2)。最初は細菌とみなされていたという経緯から、細菌を真似ている(mimic)という意味でミミウイルスと名付けられた。
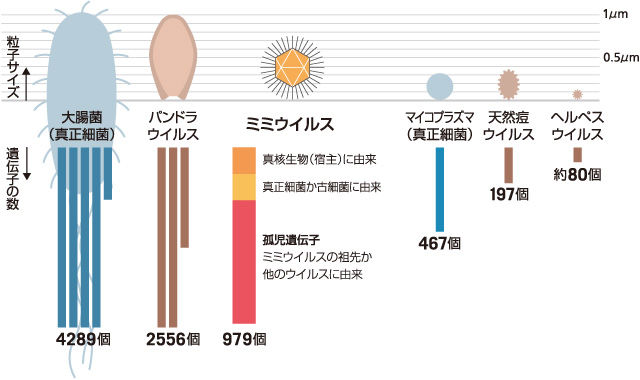
(図2)
ウイルスと真正細菌の比較。ヘルペスウイルスは従来から知られているウイルスの代表として、それ以外のウイルスは巨大ウイルスのグループとしてあげた。標準的な真正細菌の代表として大腸菌を、小型の真正細菌の代表としてマイコプラズマをあげた。
3.ミミウイルスの巨大ゲノム
2004年、私が所属するジャン=ミシェル・クラヴリらのグループとラウルトのグループによって、ミミウイルスのゲノムが解析され、それが線形の2本鎖DNAであり、サイズは約1.2Mbpもあることが分かった。粒子サイズだけでなく、ゲノムサイズも大きいのである。ちなみに、ウイルスの中でこれまで大きいとされていた天然痘ウイルスやクロレラウイルスのゲノムサイズは約0.2—0.3Mbp(天然痘ウイルスの遺伝子数は197個)、小型の真正細菌・マイコプラズマのゲノムサイズは約0.6Mbp(遺伝子数は467個)である。他にも多くのゲノムサイズを比較してみたところ、ミミウイルスは真正細菌と古細菌に匹敵する大きさだったのである。
さらに詳しく解析した結果、タンパク質をコードしている遺伝子が約1000個もあり、ミミウイルスのゲノムには遺伝子が高密度に詰まっていることが明らかになった(図2)。そのうち約40%は細胞性生物(原核生物と宿主を含む真核生物)の遺伝子と相同性を示し、残りの約60%は自身の祖先から受け継いだもの、あるいは別のウイルスから伝播したものと考えられている。この60%の部分は、既存のデータベースに類縁配列が無い孤児遺伝子である。遺伝子の系統解析の結果や、大量の孤児遺伝子を保持していることから私たちは、巨大ウイルスの由来は極めて古く、他の生きものと並ぶ新たなグループとして考えたいと思っている(図3)。

(図3)
巨大ウイルスも含めた生きもののグループ分け。由来が分からない孤児遺伝子の多さから、巨大ウイルスも生きものと並ぶ新たなグループではないかと考えている。
ところで、ミミウイルスはアミノ酸をtRNAに結合するアミノアシルtRNA合成酵素など、リボソーム以外の翻訳に関わる分子の遺伝子をもっていることが明らかになった。通常のウイルスが遺伝情報の翻訳を宿主細胞に委ねているのに対し、ミミウイルスは自己複製を完全にアメーバに頼っているわけではなく、通常のウイルスよりも高い自立性を保持していると言える。高度な遺伝情報を保持し、生物の重要な特徴である翻訳系を部分的にでも保持しているミミウイルスを、生きものではないと言い切るのは難しいのではないか。
4.宿主内に現れるウイルス粒子生産工場
環境中のミミウイルスは、アメーバの貪食作用によって細胞内に取り込まれ、食胞内に保持される(図4)。2時間ほど経つと、ミミウイルスはスターゲートと呼ばれる特徴的な5角形の星状の門を開き、アメーバの細胞質内にゲノムを放出する。他のウイルスと同様にウイルス粒子が消滅する期間を経て、約5時間後にウイルス粒子生産工場と呼ばれる巨大ウイルスに特徴的な構造をアメーバの細胞質内につくる。その大きさはアメーバの核に匹敵するほどであり、中心部ではDNAの複製が行われ、その周辺部で合成される膜とカプシド内にDNAが封入される。こうして完全体となったミミウイルスは、アメーバの細胞膜を溶解して壊し、環境中に出て行く。
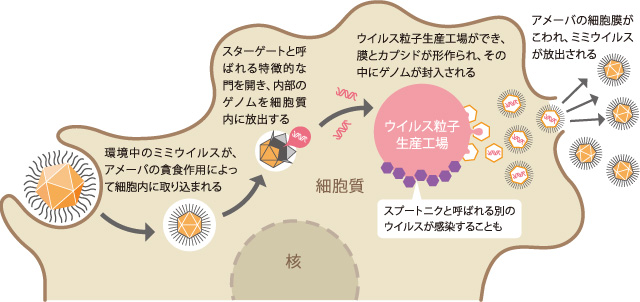
(図4)
ミミウイルスのライフサイクル。アメーバに取り込まれたミミウイルスは、アメーバの細胞質内に独自のウイルス粒子生産工場を作って増殖する。
2008年、ラウルトらによって、このウイルス粒子生産工場に感染する別のウイルスが見出された。世界初の無人人工衛星にちなんでスプートニクと名付けられたこのウイルスは、直径約50nmで約0.02Mbpの2本鎖DNAをもっている。スプートニクの特徴は、単独でアメーバに感染することはできず、必ずミミウイルスあるいはミミウイルスが作るウイルス粒子生産工場に感染することである。あたかも、このウイルス粒子生産工場が生きもののようである。私たちは、環境中にあるミミウイルスの粒子単独の状態よりも、この感染状態こそがミミウイルスの本来の姿ではないかと考えている。
5.海洋の巨大ウイルス
私たちが暮らす地球は、表面積の70%が平均4kmの深さをもつ海洋によって占められており、ウイルスも含めた海洋微生物の役割を読み解くことが、生物界全体と地球環境との関係を明らかにするうえで重要と考えられている。2009年から、私たちは世界150カ所で海水を採取し、ゲノム解析を行った結果、ミミウイルスと類似のDNA配列を多数見出した(図5)。現在のところ、ゲノムが0.3Mbpを超えるウイルスは26種ほど知られているが、今後も水圏からさらに多種の巨大ウイルスが発見されるに違いない。
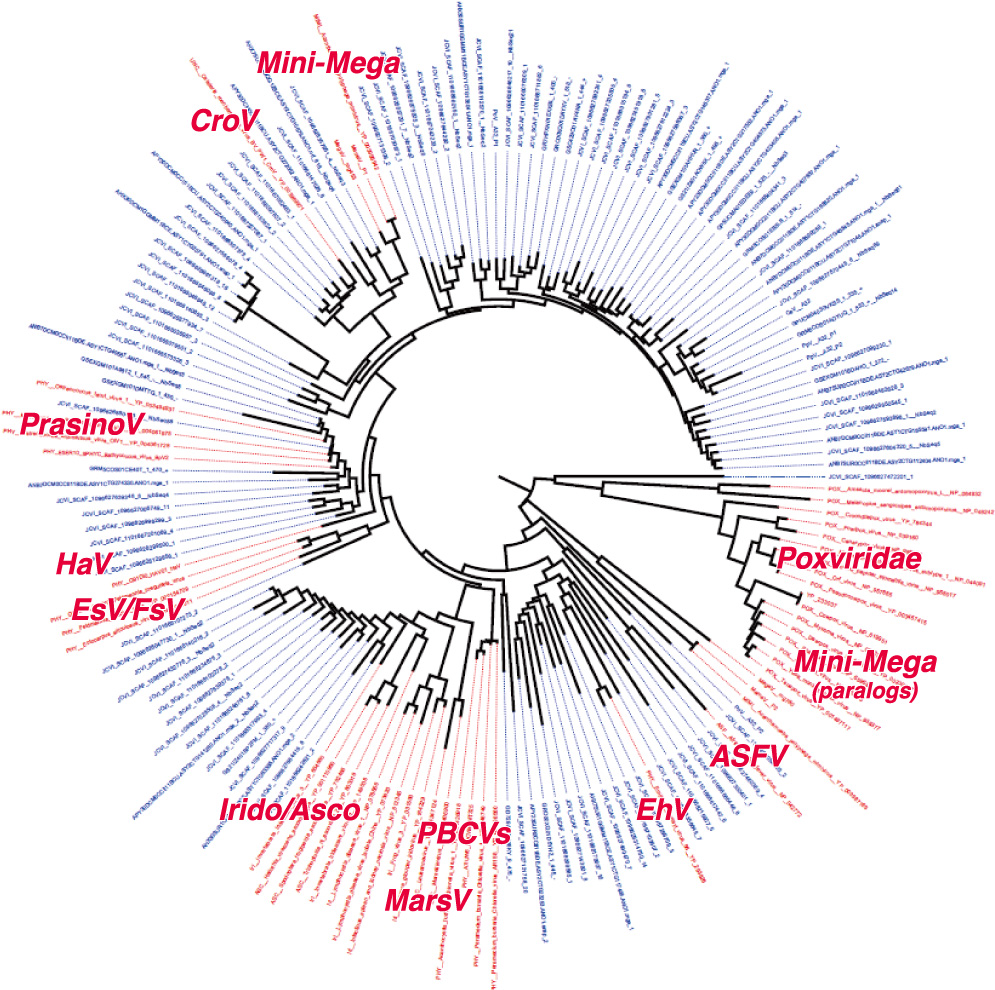
(図5)
ゲノム解析で得られた巨大ウイルスの系統樹。赤文字は既にゲノムが決まっている巨大ウイルスの系統、青文字は海水のメタゲノム解析から存在が推測される系統。
実は2013年に、長径約1μm、ゲノムサイズ2.5Mbpの超巨大なウイルスが、チリ沿岸部トゥンケン川河口とメルボルン近郊の淡水沼地から見つかっている。パンドラウイルスと名付けられたこのウイルスは、ミミウイルスも含めたこれまでのウイルスと異なり、幾何学的な構造ではなく滑らかな卵型をしていた。さらに2014年には、3万年以上前のシベリアの永久凍土から、長径約1.5μmのウイルスが見つかった。ピソウイルスと名付けられた超巨大なこのウイルスは、ゲノムサイズは0.6Mbp程度しかなかった。
私たちは今、計算科学的な手法を用い、これら巨大ウイルスの研究を進めている。彼らがどのように進化し、宿主の細胞内で何をし、そして環境中でどのような役割を担っているのか。ウイルス界を理解するに止まらず、生物界の新しい見方につながるのではないかと考えている。

緒方博之(おがた ひろゆき)
1996年 京都大学理学研究科 博士課程途中退学 1998年 博士(理学)、1996年 京都大学化学研究所 助手、1999年 仏国CNRS(国立科学研究センター)博士研究員、2001年 仏国CNRS 研究員、2008年 仏国CNRS 主任研究官、2012年 東京工業大学情報生命博士教育院 特任准教授、2014年 京都大学化学研究所 教授
.jpg)

.jpg)
.jpg)










