

RESEARCH
植物の世代を切換えるスイッチ遺伝子
動物・植物・菌類からなる多細胞真核生物は、ゲノムを1セットの状態と、2セットの状態を切換えながら子孫を残してきた。私たちは陸上植物の共通祖先から初期に分岐したコケで、その切換えに関わる遺伝子を見つけた。ここから陸上植物の進化を考えてみたい。
1.何度も起きた生物の多細胞化
樹上でさえずる鳥、足元のコケやキノコなど、私たちが眼にする生きもののほとんどは多細胞生物である。生命38億年の歴史の中で、真核生物の多細胞化は動物・植物・菌類の系統で独立に起こり、系統ごとにユニークな形や生き方をもたらした。
細胞真核生物の生活環には、細胞当たりのゲノムが1セットの「単相」と2セットの「複相」の時期があり、減数分裂と受精(接合)によって切り替わる。たとえば私たち脊椎動物は、精子と卵が受精し、受精卵が体細胞分裂して初めて多細胞の体になる。これは複相にのみ多細胞体をもつ「複相型生活環」に相当する(図1)。

(図1) 多細胞生物の生活環
シャジクモ写真提供:坂山英俊博士(神戸大学)
一方、植物と菌類は、複相だけでなく単相にも多細胞体をつくる。つまり私たちの精子や卵にあたる細胞が、受精前にそのまま分裂して多細胞化するのだ。たとえば、シャジクモなど陸上植物に近縁な藻類は単相にのみ多細胞体をつくり、複相の受精卵は多細胞化しない「単相型生活環」を示す(図1)。また、一部の藻類や陸上植物のように、単相と複相の両方で多細胞体をつくる生物もいる(図1)。この場合、単相・複相のそれぞれで独立した生活をもつことになるため、それぞれを「単相世代」・「複相世代」と呼んで区別する。その切換わりは「世代交代」と呼ぶ。私たちヒトとは異なるので不思議に見えるが、決して特異な生き方ではない。
2.世代交代から陸上植物の進化を探る
約4億7,000万年以上前、水中の藻類の一部が陸へ進出して陸上植物となって、地表を覆い、その後巨大化して森をつくった。私たちはこのような進化の背景を、世代交代のしくみから探っている。
陸上植物はすでに述べたように、単相に加えて複相でも多細胞体をつくり、世代交代をするようになった(図2)。とはいえ、初期に分岐したコケ植物は一生の大半を単相で過ごす。葉と茎をもつおなじみの姿は単相世代の多細胞体であり、複相世代につくられるのは「胞子のう」という小さな胞子の入れ物と、それを単相世代の体に癒合させる構造のみである。これらを合わせた複相世代の多細胞体全体を「胞子体」と呼ぶ。一方、コケより後に分岐した維管束植物(シダ植物と被子植物)は、単相より複相で過ごす期間が大幅に長い。被子植物の葉と茎はもちろんのこと、色鮮やかな花や甘い果実などはすべて複相世代の多細胞体、つまり胞子体である(図2)。陸上植物の歴史の中では、単相から複相へと生活環の中心が移行し、胞子体が複雑化したことが大きな変化だったと言える。
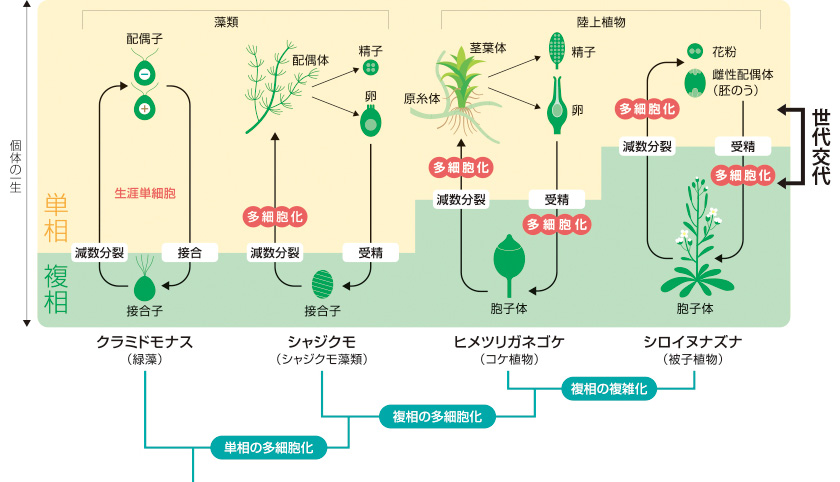
(図2) 陸上植物の生活環の進化
(註1) ホメオボックス遺伝子
DNA結合領域としてはたらくタンパク質ドメイン(ホメオドメイン)をコードするDNA配列(ホメオボックス)を持つ遺伝子の総称。多くが、形態形成、器官形成、細胞分化などの遺伝子の転写制御に関わる。
(註2) 転写因子
転写反応においてRNAポリメラーゼ以外に必要とされるタンパク質で、この場合、転写開始に関与するものを指す。
3.単細胞藻類に世代交代の原型をみる
世代交代を行う生きものには形づくりのプログラムが2通りあり、単相と複相の間でそれを切換えている。そのしくみを知ることが、世代交代の進化を探る重要な鍵となるはずだ。2008年、陸上植物と十数億年前に分岐した単細胞藻類・クラミドモナスで、世代交代の原型ともいうべき切換えのしくみが発見された。
クラミドモナスの単相の細胞は、栄養が欠乏すると配偶子へと分化する(図3)。配偶子はプラスとマイナスのいずれかの交配型を示し、異なる交配型間で接合する。このとき、プラス型配偶子にはBELL遺伝子、マイナス型配偶子にはKNOX遺伝子というそれぞれ独自のホメオボックス遺伝子(註1)がはたらいている。2つの遺伝子がつくるタンパク質は、接合によって出会うと複合体となって核へ移動し、核ゲノムに作用して複相世代特異的な遺伝子をはたらかせる転写因子(註2)となる。つまり複相への切換えスイッチの役割を果たすことがわかった(図3)。このしくみを発見したLee博士らは、BELL・KNOX遺伝子が陸上植物に受け継がれ、世代交代に重要な役割を果たすようになったのではないかという仮説を提唱した。
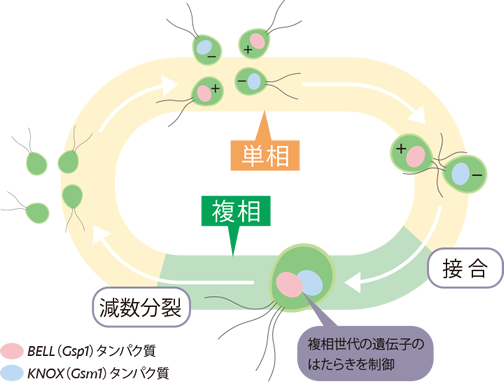
(図3) クラミドモナスのBELL遺伝子・KNOX遺伝子による相の切換え
Lee et al., Cell, 133(5),(2008) Fig.1より改変 http://dx.doi.org/10.1016/j.cell.2008.04.028
4.BELL遺伝子とKNOX2遺伝子は陸上植物の世代交代を制御する
そこで陸上植物を調べたところ、BELL遺伝子とKNOX遺伝子は陸上植物に受け継がれる過程で、遺伝子重複(註3)によって数を増やしていることがわかった。KNOX遺伝子は、陸上植物の出現に先立って、KNOX1とKNOX2という機能の異なる遺伝子群に分かれたことも明らかにされた。ヒメツリガネゴケの形づくりの遺伝子について調べていた私たちは、このKNOX2遺伝子群がコケの世代交代にはたらいていることを発見したので、それを詳しく紹介しよう。
まず、KNOX2遺伝子群をゲノム中から取り除いた株を育て、単相世代から複相世代への切換わりの変化を観察した。通常、コケの精子と卵が受精して複相世代に入ると、受精卵が多細胞化して胞子体が形成されるのだが(図4)、KNOX2がない株では胞子体の形成が初期段階で止まった。そして、胚から原糸体によく似た突起が出て来たのである(図4)。胚を取り出して育ててみると、この突起から単相世代の姿である茎葉体が育った(図4)。ここから、KNOX2は複相世代に単相世代特有の形づくりが行われないよう抑制するはたらきをしていることがわかる。
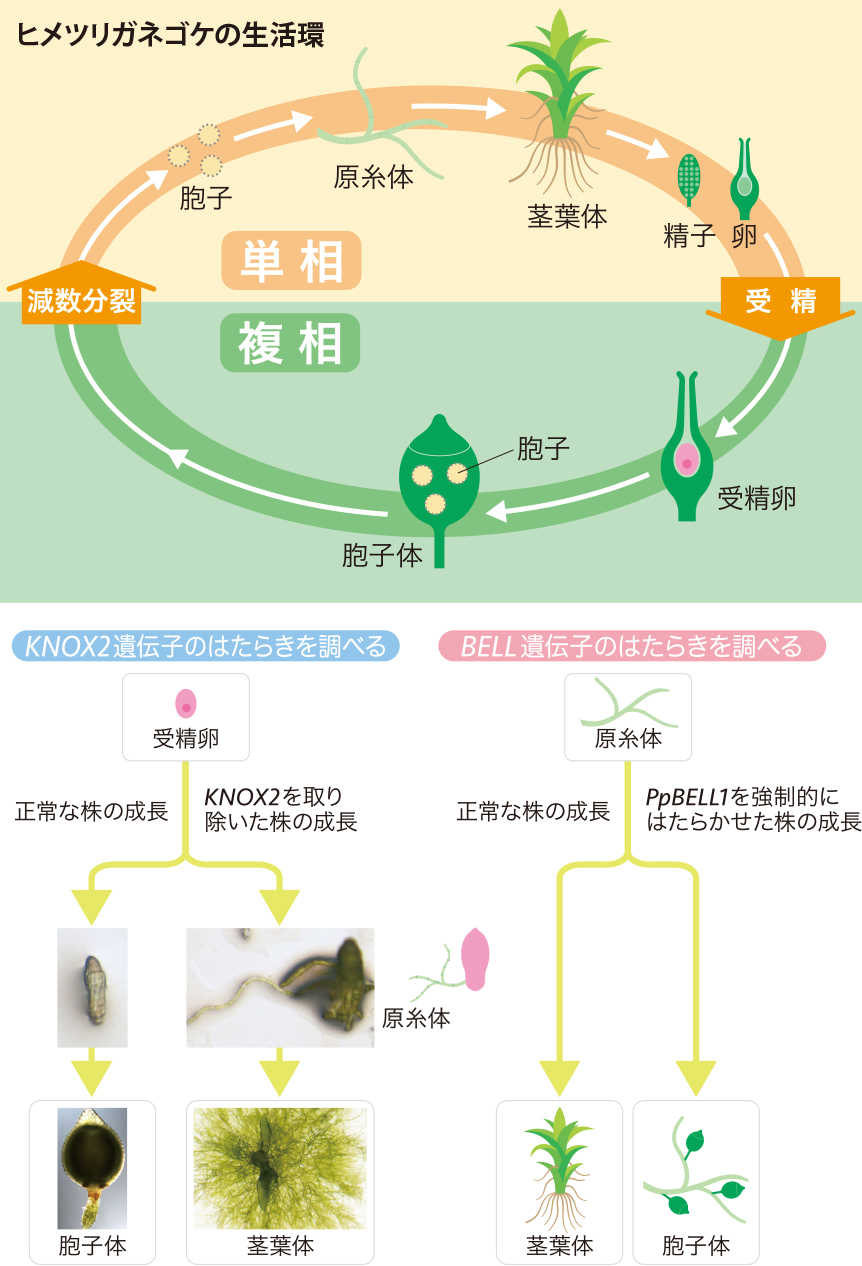
(図4) ヒメツリガネゴケのBELL遺伝子とKNOX2遺伝子のはたらき
PpBell1の結果については、Horst et al., Nature Plants 2, 15209 (2016) に基づく
一方、BELL遺伝子も世代交代にはたらいていることを、2016年にHorst博士らが明らかにしている。ヒメツリガネゴケのBELL遺伝子の一つ、PpBELL1遺伝子を単相世代で強制的にはたらかせると、複相世代の姿である胞子体が単相世代の原糸体上に形成されたのである(図4)。つまり、この遺伝子が複相世代の形づくりを開始する転写因子だったことになる。こうしてPpBELL1遺伝子が複相世代の形づくりを開始し、KNOX2遺伝子が単相世代の形づくりを抑制することによって、単相世代から複相世代への切換えが起きるというしくみが見えてきた(図4)。
さらに私たちは、陸上植物の出現に先立って分かれたKNOX1遺伝子群についても調べ、それが細胞の増殖を促すことを明らかにした。PpBELL1とKNOX2が世代の切換えを行い、さらにKNOX1が胞子体の体作りに重要な分裂組織ではたらくことで、複相世代特有の多細胞体である胞子体がつくられるのだろう。コケ植物は、陸上植物の共通祖先から初期に分岐したグループであり、祖先から受け継いだ切換えスイッチのはたらきに新たな機能を加えて、多細胞体をつくるようになったという進化の姿を見せてくれているのである。
(註3) 遺伝子重複
染色体分裂時のDNA不等乗り換えやトランスポゾンなどによって、ゲノム内で同一の遺伝子が複数存在する状態になること。生物進化に伴う遺伝子数の増加や新しい機能の獲得に重要な役割を果たしていると考えられている。
5.陸上植物の複雑化に貢献したBELL遺伝子とKNOX遺伝子
被子植物についても、BELL遺伝子とKNOX遺伝子のはたらきが明らかになってきている。シロイヌナズナのKNOX1遺伝子は、細胞の増殖を促すという性質を受け継いでいる。一方、シロイヌナズナのKNOX2遺伝子をはたらかないようにすると葉が切れ込む表現型を示すことから、この遺伝子には細胞の分化を促すはたらきがあると考えられる。シロイヌナズナでは、KNOX1遺伝子とKNOX2遺伝子は、協調して複相世代の複雑な形づくりにはたらいているのだろう。
シロイヌナズナでは、BELL遺伝子が遺伝子重複によって増えており、少なくともそのうちのいくつかはKNOX1遺伝子あるいはKNOX2遺伝子とともにはたらくことが示唆されている。KNOX1・KNOX2遺伝子の出現と、BELL遺伝子の重複とが機能の分化を促し、陸上植物の複相の長期化に貢献したのであろうと考えられる(図5)。
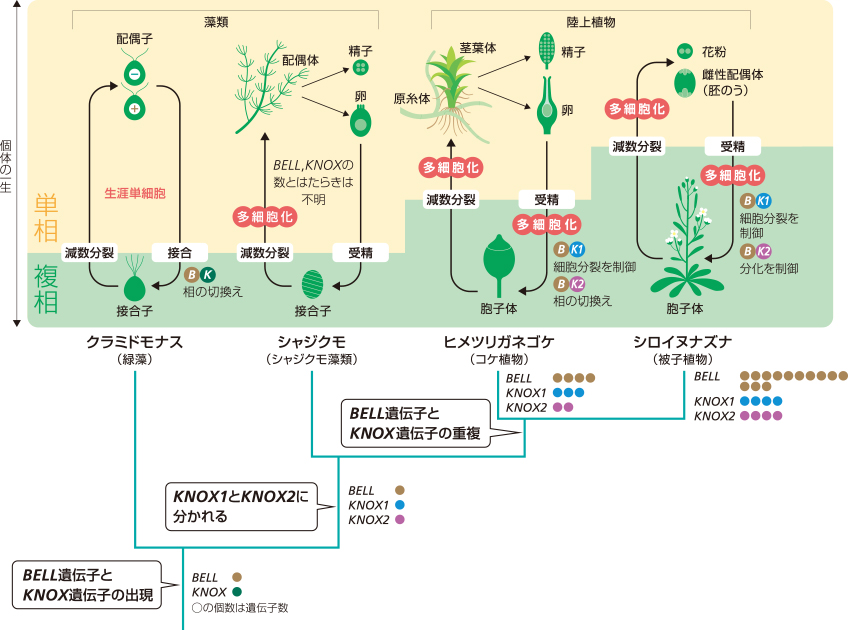
(図5) BELLとKNOXの遺伝子重複と生活環の進化
6.世代交代の共通性は?
近年、多細胞動物に近縁な単細胞動物のゲノム解析が行われ、動物は形づくりの遺伝子の大半を多細胞化以前に獲得していることがわかった。ここから、動物では同じ遺伝子のはたらき方の違いが形の違いをつくり出すのに重要だと考えられる。一方植物では、形づくりに関わる遺伝子が種によって異なっている場合がある。植物は、進化における形と遺伝子の関係を解くよい実験場なのかもしれない。
私たちは主として陸上植物での世代交代を研究してきたが、世代交代は陸上植物のほかにも紅藻類や褐藻類などでもみられる(図6)。これら起源の異なる世代交代がどのように制御されているのかも、とても興味深いところだ。近年褐藻類のシオミドロでも、複相世代に単相の形がつくられる変異体(註4)がみつかっているが、その原因遺伝子はまだわかっていない。BELL遺伝子とKNOX遺伝子は紅藻と緑色植物でしか確認されていない。シオミドロではどのような遺伝子が世代交代にはたらいているのだろう。異なる系統で生み出された世代交代に、共通性はあるのだろうか。今後、褐藻を含めさまざまな系統の生物で、世代交代を制御するスイッチの解析が進み、世代交代に関する情報が共有されることで、真核多細胞生物の進化の全体像が明らかとなるだろう。
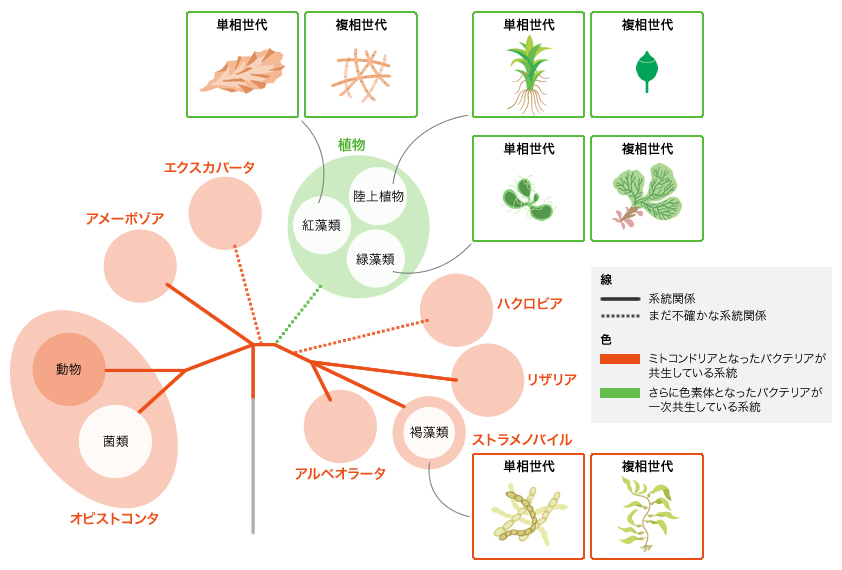
(図6) 世代交代は独立に進化した
白丸で示したグループに、世代交代を行う生物がみられる。紅藻類・緑藻類では一部の種が、褐藻類では多くの種が世代交代を行う。また菌類では、担子菌類、子嚢菌類、卵菌類などの一部の種に世代交代を行うものが見られる。
(註4) 変異体
突然変異体、突然変異株ともいう。ゲノムDNAの配列に何らかの突然変異を生じた生物、細胞またはウイルスのこと。

榊原 恵子(さかきばら・けいこ)
2003年総合研究大学院大学にて博士(理学)取得。山口県立萩高等学校教諭(臨時)、日本学術振興会特別研究員(PD)、オーストラリア・モナシュ大学博士研究員、科学技術振興機構ERATO長谷部分化全能性プロジェクト技術参事、東京大学大学院理学研究科助教などを経て、2016年より立教大学理学部准教授。
.jpg)

.jpg)
.jpg)










