

Special Story
共生・共進化 時間と空間の中で
つながる生きものたち
種の異なる生きものが相互に何らかの関係を結びながら生息するのが共生ですが,それでは,種とはいったい何でしょう?お互いに種の独立性を維持しながら共存する仕組みがあるはずです。ここでは,シロイヌナズナを用いて,雌が雄との交配を拒否する仕組みの研究を紹介します。この細胞レベルの研究を,DNAレベルの研究に進めたのは,じつはフィールドの調査がきっかけ。雌雄間異種認識機構の研究がDNA からフィールドまでを貫いて,共生の仕組みを解くキーになるかもしれません。
地球上には100万種をゆうに超える多様な生物が共生していてさまざまな関係をもつが,そのうちでも,有性生殖過程で雌と雄が出会う場面は重要である。とくにここで,他種の異性に注目すると,そもそも種とは何かという大きな問いが見えてくる。現在の種概念の鍵は,生殖隔離という,他種との交配を防ぐ仕組みだからである。ヒトとチンパンジーが別種なのは,ヒトが直立二足歩行だからでも,ましてやヒトのほうが頭がいいからでもなく,ヒトとチンパンジーの間には子孫が育たないからだ。この種概念は20 世紀半ばに確立したが,生殖隔離の仕組みの実証データは未だ乏しい。野外では多数の種で同種や他種の異性との出会いが起きているはずだ。ここで,交配相手を選ぶ過程を植物を通して見ていこう。
実験材料としてはシロイヌナズナとその近縁種を用いる。シロイヌナズナはアブラナ科の草本で,2000 年末に日米欧の協力で全ゲノム配列が解読され,研究のための環境が整ったモデル生物だ。
まずシロイヌナズナの雌しべに,近縁種であるハナナ(ムラサキダイコン)の花粉をかけあわせたところ,花粉管(雄細胞)は,雌(雌性配偶体)へ向かって伸び始めたが,途中で方向を失って迷走し,受精に至らなかった。このような迷走は,すでに雌性配偶体に異常があるシロイヌナズナ(magatama 変異体)同士の雌雄間相互作用で観察していた。雄は雌性配偶体の近くにまで誘引されるが,直前で迷走してしまうのだ(図)。おそらくハナナの花粉管はシロイヌナズナの雌性配偶体からの誘引シグナルを認識できないのだろう。ナズナ(ぺんぺん草)の花粉を用いると,花粉管がすぐに伸びなくなり,タネツケバナでは届いても精細胞を放出できない。ここから,有性生殖は,雌雄間での何段階にもわたる認識の連続であること,異種の雄は,どこかの段階でシグナル交換に失敗して先へ進めなくなることがわかった。これは動物でも同じであり,適切な配偶相手を選び出す巧みな仕組みの一つと考えられる。雌がちゃんと呼んでやらないと雄がさまようのは,植物だけではないようだ。
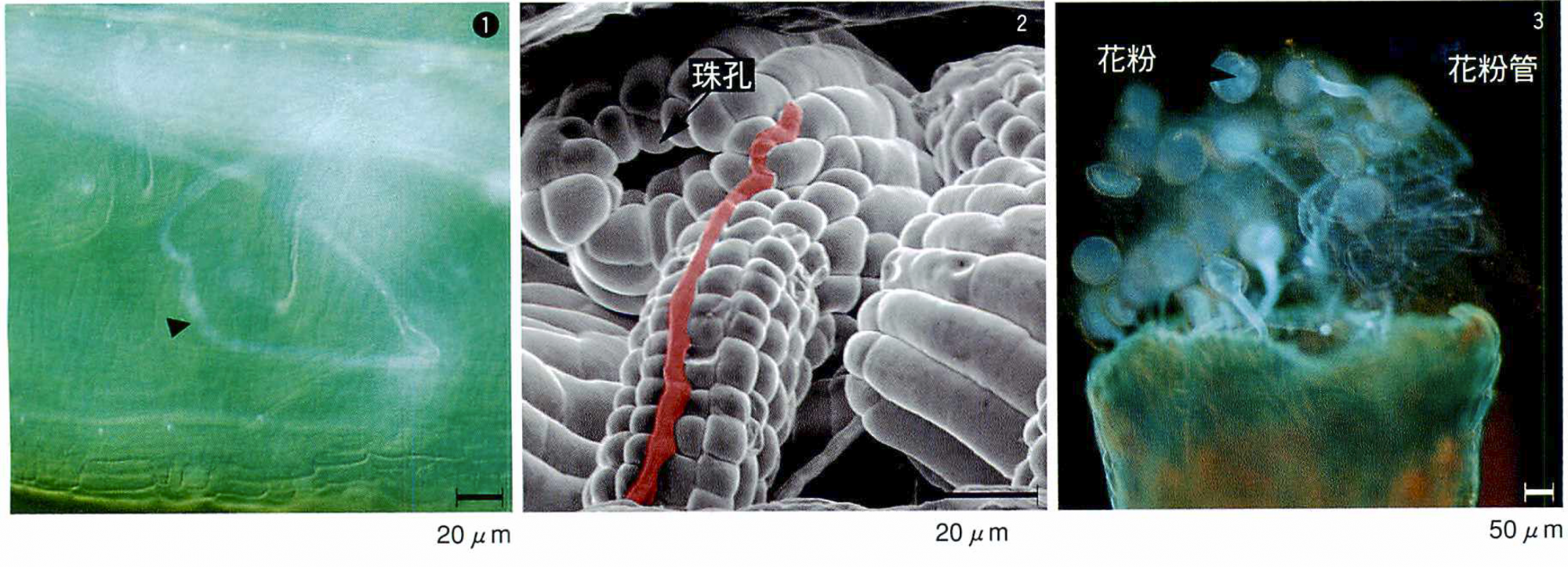
①シロイヌナズナの雌(雌性配偶体)を見つけられず迷走したハナナの雄(花粉管)。
②雌性配偶体の発生が異常なシロイヌナズナmagatama 突然変異体における花粉管の道筋(赤い線/電子顕微鏡写真)。花粉管は,珠孔の中にいる雌性配偶体のすぐ近くまで伸びたが,結局別の方向へ。
③フタバガキの自家不和合性の機構。同じ個体の雌しべと花粉をかけあわせると,花粉から花粉管は発芽するが,雌しべの先端で糸球のようになって迷走。花粉管がこの段階から先へ進めないことが,近親交配を防ぐ。
有性生殖で拒むのは,何も他種ばかりではない。近親交配では子孫が弱るので,雌雄両性の植物は,自分自身の雌と雄の受精を防ぐ自家不和合性という仕組みを発達させていることが多い。マレーシア熱帯林冠の主要樹種であるフタバガキも自家不和合性なので,この仕組みを生態学者の田中健太氏(京都大学生態学研究センター)と一緒にマレーシアで調べたところ,フタバガキの自家不和合性は花粉管の迷走によることがわかった。これは,雄が迷うことによる自家不和合性のはじめての報告である。不適切な雄が排除されるシステムは,種間だけでなく種内でも見られるのだ。
これからは,こうした細胞レベルから,種や個体間の違いの起因となるDNA 塩基配列レベルの研究に進みたいと思っていたところ,ひょんなことから研究の手がかりが見えてきた。山歩きをしていたら,たまたまシロイヌナズナに似た花を見つけた。これがミヤマハタザオという植物で,後にシロイヌナズナに最も近縁な植物の一つであることが明らかになった。シロイヌナズナと交配してみたところ雑種が育ち,雑種の花粉や卵細胞は育たなかった。つまり子は育つが孫が育たないという生殖隔離がみられた。重要なのは雑種が一代でも育つことで,これによってシロイヌナズナの膨大な知見を活かした遺伝学的な研究ができる。たとえば,シロイヌナズナのGL1 遺伝子を欠損して葉面の毛がなくなった変異体と,葉面の毛がないミヤマハタザオの系統をかけあわせたところ,その雑種には毛がなかった。そこでミヤマハタザオのGL1 遺伝子を調べると,1 塩基挿入や欠失が起こったために遺伝子が壊れ,毛を失ったことがわかった。

この延長で研究を進めていけば,雌雄の相互作用を担う遺伝子群のここが違うために種が違うのだ,といった研究もできる。動物では,モデル生物ショウジョウバエとその近縁種を用いた研究から,生殖隔離に関わる遺伝子の候補が見つかっている。種というものが,聖書で神が創造したような確固とした不変のものでないということは,ダーウィンが19 世紀に述べている。雌と雄がもつ膨大なゲノムのうちのごく一部が違っているために協調できず,種が違ってくるのだということが明らかになる日も遠くなかろう。
生物多様性を分子・細胞レベルから理解するには,分子遺伝学者とフィールド研究者との協力が不可欠だ。藤井伸二氏(大阪市自然史博物館)に連れられて琵琶湖や伊勢湾でミヤマハタザオの低地型であるタチスズシロソウを調査したとき,分類学者の膨大な知識に驚かされた。本来数十年に一度の洪水でできる砂地に暮らし,他の植物が移入する頃には次の洪水でできた砂地で育つという興味深い生活様式をもつタチスズシロソウは,水辺が人工化したために絶滅危惧種となってしまったとのこと。墓地の砂地などで細々と生きているらしい。このような自然史の知識から解析可能なものを拾い出し,モデル生物の遺伝学を活かせば,生物の多様性を,生命システムの多様性として統一的に説明できるだろう。種や多様性は一筋縄ではいかない問題だが,DNA の実証的データを積み重ね,実験室とフィールドをつなぐことに挑戦していくつもりだ。

⑤ハナナ。
鴨川(京都)にて。

⑥ミヤマハタザオの低地型であるタチスズシロソウ。琵琶湖岸の墓地の砂上にひっそりと生きている。
⑦ハクサンハタザオの一面の草原。ミヤマハタザオと同様に,シロイヌナズナとの雑種ができる。大阪府多田銀山跡にて。
清水健太郎(しみず・けんたろう)
1974 年埼玉県生まれ。京都大学理学研究科博士課程在籍。分子生物学を用いたシロイヌナズナ研究の創始者の一人,岡田清孝教授の指導のもと博士論文執筆中。雌雄間相互作用を中心に,植物の多様性と進化を研究。数学者広中平祐の創設した「数理の翼セミナー」と「湧源クラブ」を中心に高校生への理科教育にも携わる。共著書に『新版植物の形を決める分子機構』(秀潤社)がある。
.jpg)

.jpg)
.jpg)










.png)