Publication List
Su Z.-H., Sasaki, A. Kusumi, J. Chou P.-A., Tzeng H.-Y., Li H.-Q., and Yu H. (2022)
Pollinator sharing, copollination, and speciation by host shifting among six closely related dioecious fig species
Commun. Biol. 5, Article number 284
The obligate pollination mutualism between figs (Ficus, Moraceae) and pollinator wasps (Agaonidae, Hymenoptera) is a classic example of cospeciation. However, examples of phylogenetic incongruencies between figs and their pollinators suggest that pollinators may speciate by host shifting. To investigate the mechanism of speciation by host shifting, we examined the phylogenetic relationships and population genetic structures of six closely related fig species and their pollinators from southern China and Taiwan-Ryukyu islands using various molecular markers. The results revealed 1) an extraordinary case of pollinator sharing, in which five distinct fig species share a single pollinator species in southern China; 2) two types of copollination, namely, sympatric copollination by pollinator duplication or pollinator migration, and allopatric copollination by host migration and new pollinator acquisition; 3) fig species from southern China have colonized Taiwan repeatedly and one of these events has been followed by host shifting, reestablishment of host specificity, and pollinator speciation, in order. Based on our results, we propose a model for pollinator speciation by host shifting in which the reestablishment of host-specificity plays a central role in the speciation process. These findings provide important insights into understanding the mechanisms underlying pollinator speciation and host specificity in obligate pollination mutualism.
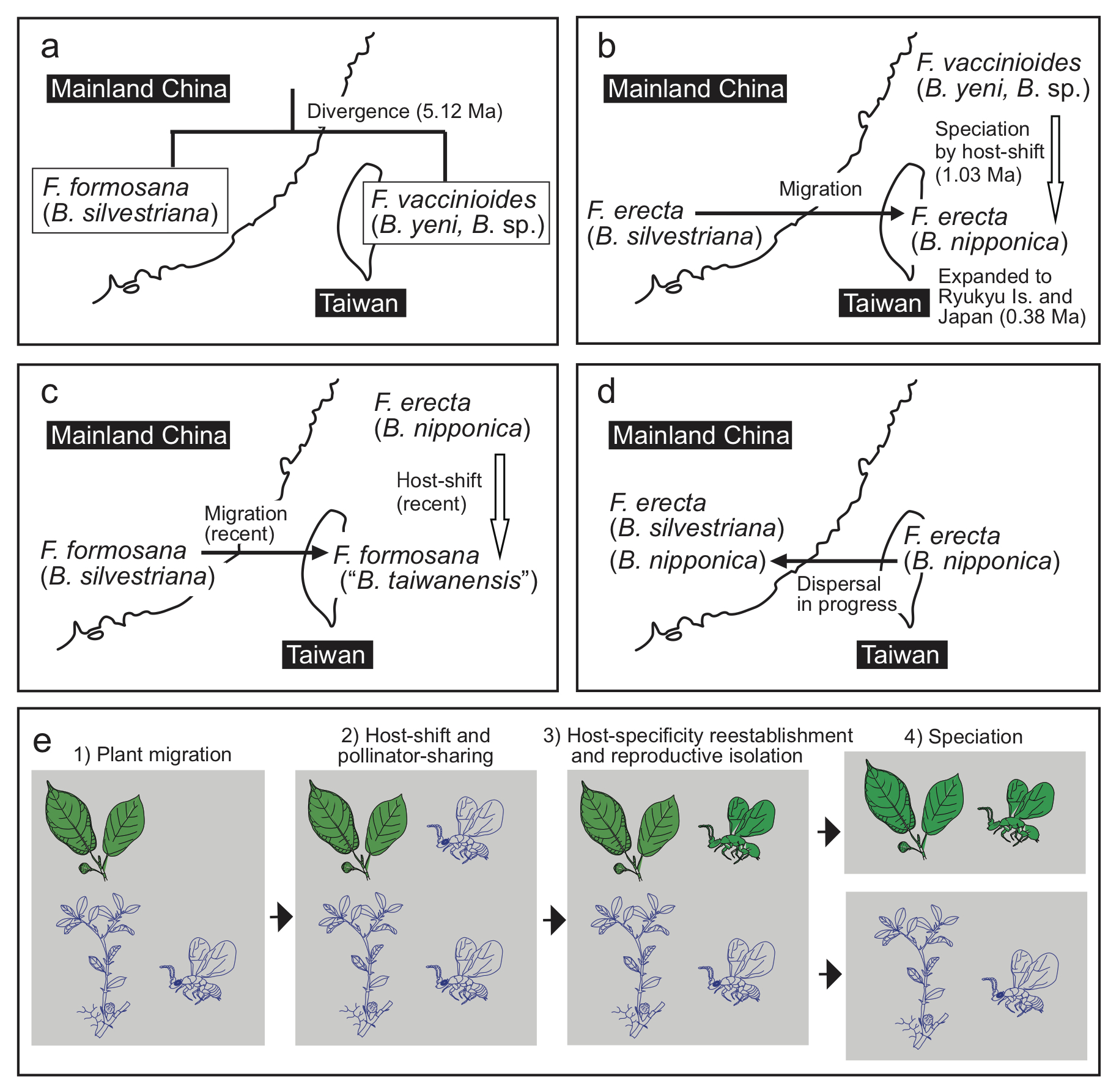 Establishment of the mutualistic relationships in the F. erecta complex and its pollinators (in parentheses) in the Taiwan-Ryukyu islands (a–d) and a model for host-shift speciation proposed in this study (e). a The divergence of F. vaccinioides–B. yeni/B. sp. and F. formosana–B. silvestriana. b F. erecta migrated into Taiwan from mainland China. Blastophaga sp. host-shifted to F. erecta and speciated into B. nipponica. Then, the distribution range of F. erecta–B. nipponica expanded northward to the Ryukyu Islands and Japan. The chronology in a and b is based on the dating analysis of the pollinators (Supplementary Fig. 10). c F. formosana probably migrated into Taiwan recently from mainland China, and B. nipponica host-shifted to F. formosana; at present, F. erecta and F. formosana appear to share B. nipponica including “B. taiwanensis” in Taiwan. d B. nipponica dispersed from Taiwan to the Fujian region (FJ) of mainland China, resulting in an example of sympatric co-pollination. e The proposed model for the host-shift speciation events described in this study consists of four stages: (1) Host figs migrate into a new niche without their original pollinators; (2) The migrated host figs are pollinated by new wasps (host shifting); as a result, two host fig species share one wasp pollinator (pollinator sharing); (3) The host-shifted wasps evolve and become associated with the host figs that migrated (host-specificity reestablishment), and reproductive isolation occurs; (4) Genetic divergence proceeds and speciation occurs. These images show only the evolutionary events, not the details of the evolution of figs and pollinators during these processes.-1
Establishment of the mutualistic relationships in the F. erecta complex and its pollinators (in parentheses) in the Taiwan-Ryukyu islands (a–d) and a model for host-shift speciation proposed in this study (e). a The divergence of F. vaccinioides–B. yeni/B. sp. and F. formosana–B. silvestriana. b F. erecta migrated into Taiwan from mainland China. Blastophaga sp. host-shifted to F. erecta and speciated into B. nipponica. Then, the distribution range of F. erecta–B. nipponica expanded northward to the Ryukyu Islands and Japan. The chronology in a and b is based on the dating analysis of the pollinators (Supplementary Fig. 10). c F. formosana probably migrated into Taiwan recently from mainland China, and B. nipponica host-shifted to F. formosana; at present, F. erecta and F. formosana appear to share B. nipponica including “B. taiwanensis” in Taiwan. d B. nipponica dispersed from Taiwan to the Fujian region (FJ) of mainland China, resulting in an example of sympatric co-pollination. e The proposed model for the host-shift speciation events described in this study consists of four stages: (1) Host figs migrate into a new niche without their original pollinators; (2) The migrated host figs are pollinated by new wasps (host shifting); as a result, two host fig species share one wasp pollinator (pollinator sharing); (3) The host-shifted wasps evolve and become associated with the host figs that migrated (host-specificity reestablishment), and reproductive isolation occurs; (4) Genetic divergence proceeds and speciation occurs. These images show only the evolutionary events, not the details of the evolution of figs and pollinators during these processes.-1