年度別活動報告
年度別活動報告書:2009年度
分子系統から生物進化を探る 3-1.イチジク属植物とイチジクコバチとの共生関係と共進化
蘇 智慧(主任研究員) 楠見淳子(奨励研究員)
佐々木綾子、尾川武史(研究補助員) 石渡啓介、宮澤秀幸、上田千晶、長久保麻子、坂内和洋(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの相利共生関係の構築、維持、崩壊のメカニズム、種分化の様式などについて分子系統解析や集団遺伝学的解析などにより研究を行っている。
イチジク属Ficusは、クワ科 (Moraceae) に属し、6亜属 (イチジク亜属Ficus, アコウ亜属Urostigma, Pharmacosycea, Sycomorus, Synoecia, Sycidium)に分けられ、東南アジア、アフリカ、中南米など熱帯を中心に約750種が世界中に分布している。日本はイチジク属の分布域の北限にあたり、南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。そのうち、小笠原諸島に固有種3種と移入種1種が生息している。一方、イチジク属植物に共生、或いは寄生している一群のイチジクコバチの仲間は、分類学的に膜翅目のコバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとされている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つが送粉コバチ亜科Agaoninae (fig-pollinating wasps)で、他の亜科はすべて非送粉コバチによって構成されている。送粉コバチ類は同定されているものだけで20属300種以上あり、植物と同様に種多様性が高い。
イチジク属植物と送粉コバチとの間、子孫を残す共通の目的のもとで、「1種対1種」関係が結ばれていると言われている。この「1種対1種」関係を維持しながら種分化が起きるとしたら、同調した種分化や系統分化が起こることが予想される。これまで、分子系統学的解析を用いてこの仮説を検証する研究が行われてきた。その結果、イチジク属の節(section)レベルの系統関係とコバチ類の属のそれとがおおまかに一致し、仮説が支持されるものの1)、種間、種内レベルでは系統関係の矛盾のほかに、送粉コバチの隠蔽種や1種のイチジク属植物に複数種の送粉コバチが共生するなど、不明瞭な点が残っている2, 3)。これまでの我々の日本産、雲南産、メキシコ産のイチジク属植物とイチジクコバチの分子系統解析からも同様の結果が示されている4)。日本産のイチジク属植物とイチジクコバチでは、それらの分子系統樹のトポロジーが一致し、「1種対1種」の関係がほぼ厳密に維持されていることが示唆された一方で、メキシコ産、中国産の材料では、低次の分類群(種レベル)の系統関係が両者で一致しない場合が見られた。これは、メキシコや雲南では、近縁種間で「1種対1種」の関係と同調的種分化の乱れや宿主転換がある程度おきていることを示している。このような共生関係の厳密性の違いは、なぜ、どのようにして生じるのだろうか?この問いに答えるべく、我々は、それぞれの地域における種間、種内レベルの遺伝的背景の詳細な解析を進めている。その答えは「1種対1種」の維持、崩壊のメカニズム、そして共種分化のプロセスを知る重要な手がかりとなりうる。
本年度、我々は、(1)日本、台湾、中国に広く分布するイヌビワ(Ficus erecta)とそれに共生、寄生するコバチ類の集団遺伝学的解析を行った。イヌビワと送粉コバチ(イヌビワコバチBlastophaga nipponica)、寄生コバチ(イヌビワオナガコバチ Sycoscapter inubiwae)を、日本の5地域及び台湾から採集し、種内の遺伝的多様性、地域集団間の遺伝的分化の程度の解析を行い、種内レベルでも同調性が維持されているか検証を試みた。また、(2)小笠原諸島固有種であるトキワイヌビワ(F. boninsimae)、オオトキワイヌビワ(F. nishimurae)、オオヤマイチジク(F. iidaiana)とそれに共生するコバチ類(Blastophaga sp.)の起源ついても新たな知見が得られたのでここに報告する。
結果と考察
1)イヌビワとそれに共生、寄生するコバチ類の種内レベルの遺伝的多様性、集団間の遺伝的分化の解析
本研究では、日本の5地域(関西、福岡、鹿児島、沖縄、八重山)と台湾で採集を行った(図1)。各植物個体の葉および花嚢を採集し、葉の一部を用いてDNAを抽出した。コバチ類は、植物1個体あたり、1~4個の花嚢から採取した。同じ親由来の個体が重複するのを避けるため、花嚢1つあたり1匹の雌を用いることとし、1個体ごとにDNAを抽出した。日本の集団に関しては、統計的解析に必要な数のサンプルを回収した(表1)。
 |
<マイクロサテライトマーカーを用いたイヌビワの遺伝的構造の解析>
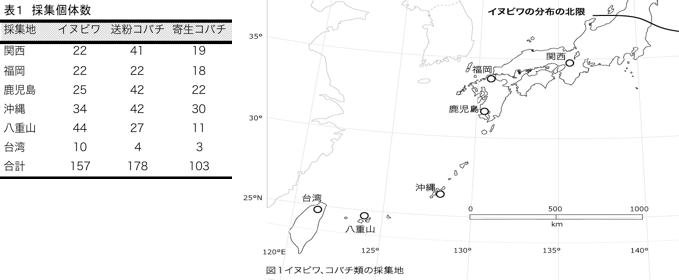
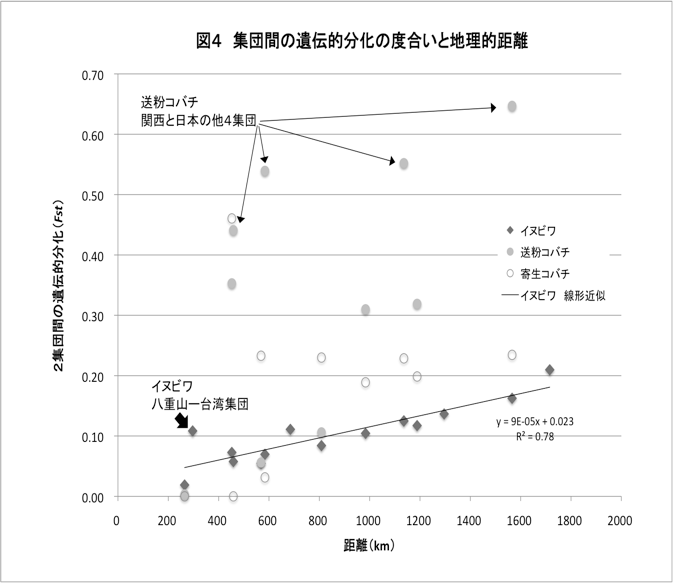
予備実験から、イヌビワの塩基配列多型は非常に少ないことが示されていたため、集団間の分化の有無を解析するには、より多型性の高いマイクロサテライトマーカーを用いる必要があった。前年度までに7つのマイクロサテライトマーカーを作成したが、本年度は新たに3つのマーカーを作成し、計10個のマーカーを用いてマイクロサテライト解析を行った。このうち1つのマーカーは日本のサンプルでの多型性が非常に低かったので、以後のデータ解析からは除外している。 マイクロサテライトデータの解析は、GeneAlEx6.3とArlequin ver 3.11というソフトウエアを用いて行った。遺伝的多様度を示すパラメーターとなる、ヘテロ接合頻度(He)やハプロタイプ数(Ne)等の値は沖縄集団が最も高く、関西集団が最も低い値を示しており、緯度に沿ったクラインがみられた(図2)。また、台湾、沖縄、八重山集団には固有のハプロタイプが多い一方で、関西の集団にはそれが全くないことを考慮すると、地域集団間の移入、移出に偏りがある可能性を示している。今後、最尤法等を用い、集団間の移住率の推定を行い、イヌビワの分布の広がりや分散能力について検討したい。 また、集団間の遺伝的分化の程度を示す統計量(Fst)を計算したところ、地域集団間の遺伝的分化があることが示された。さらに、2つの地域集団間のFstとその地理的距離には有意な相関が見られた(Mantel test p<0.01)(図4)。従って、イヌビワの遺伝子流動(花粉や種の移動)は地理的距離が大きいほど制限される度合いが強いことが予想された。ただ、台湾と八重山集団は地理的距離が短いにも関わらずFstの値が高くなっており、地理的距離だけでは説明できない場合も見られた。
 |
 |
<塩基配列多型を用いたイヌビワの送粉コバチ、寄生コバチの遺伝的構造の解析>
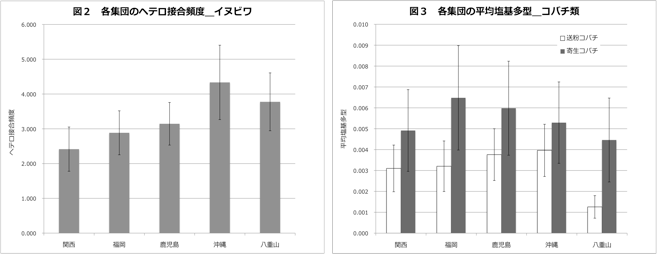
送粉コバチについては、ミトコンドリアのCOI, COII(1909 bp) CytB(432 bp)遺伝子、寄生コバチについては、同じくCOI(868 bp) CytB(432 bp)遺伝子の塩基配列多型を用いて解析を行った。データの解析には、Arlequin ver 3.11とDnasp 5.0ソフトウエアを用いた。塩基多様度、平均塩基多型を比較したところ、全集団において寄生コバチが高い値を示したものの、有意な差がみられたのは八重山集団のみであった(図3)。八重山集団では、特に送粉コバチの塩基多様度、平均塩基多型が低く、他集団に比べ集団サイズが小さいと考えられる。
イヌビワと同様に、それぞれのFstを計算したところ、どちらのコバチも集団間の遺伝的分化があることが示された。しかし、地理的距離との相関は見られなかった(図4)。両方のコバチの特徴としては、台湾集団と日本の5集団で共有するハプロタイプが無く、ハプロタイプ間の変異の蓄積も非常に多かった。送粉コバチの場合、台湾―日本集団間の変異量は、台湾の近縁種間の変異量を超えており、これは、台湾と日本との間でコバチの移入、移出が断たれてから長い時間が経過していることを示唆している。
また、送粉コバチの特徴としては、関西集団で固有のハプロタイプの頻度が非常に高くなっており他の日本集団との極端な遺伝的分化が示された(図4)。このような特徴は、イヌビワや寄生コバチにはなく、関西の送粉コバチが何処に由来するかは非常に興味深い。
<イヌビワ、送粉コバチ、寄生コバチの集団間分化の程度の比較>
他集団との間に、極端に高いFstの値がみられた台湾集団と関西集団を除き、3つの生物それぞれについてAMOVA解析(分散分析)を行った。遺伝的変異量の全分散に対する集団間の分散の割合はコバチ類でより高い値を示した。(イヌビワ5%、送粉コバチ18%、寄生コバチ22%)。これは、コバチ類では集団間の分化の程度が高いことを示しており、コバチ類の移動能力が非常に限られていることが要因の一つであると考えられる。イヌビワは、花粉の移動はコバチに依存しているものの、鳥や小動物を介した種子による移動があるため、広い範囲に拡散できると考えられる。
2)小笠原諸島固有イチジク属植物とそれに共生するコバチ類の起源
2-1. イヌビワのイヌビワコバチ受け入れ期間の延長とその仕組み
典型的な海洋島である小笠原諸島には、大陸から分布を拡大した生物が島内で独自に進化を遂げた固有種が多くみられ、イチジク属も3種の固有種(トキワイヌビワ、オオトキワイヌビワ、オオヤマイチジク)が生息している。これらは、単一の祖先が島内で種分化したものと考えられているが、その起源は不明であった。そこで、核と葉緑体DNAの塩基配列を用い、日本、台湾に分布する近縁種を含めた分子系統解析を行った。葉緑体DNAの塩基配列に基づいた分子系統学的解析結果では、日本に分布するイヌビワが固有種の祖先系統に最も近いことが示された。ところが、核遺伝子(G3pdh Aco1)、遺伝子間領域(ITS)では、イヌビワと固有種は姉妹群を形成しない場合が見られた。葉緑体と核遺伝子との結果の相違は、固有種の祖先系統でイヌビワを片親とする雑種形成が起こった可能性を示唆している。また、イチジク属植物と共生関係にあり、共種分化しているとされるコバチ類の分子系統解析も行った。解析には、ミトコンドリア遺伝子(CytB)、核遺伝子 (28SrRNA) を用いた。その結果、小笠原のイチジク属固有種と共生するコバチはイヌビワのコバチと姉妹群を形成せず、その祖先系統は別種のコバチに由来する可能性が高いことが示された。本研究では、イチジク属固有種とそれに共生するコバチの由来を特定するには至らなかったが、小笠原諸島のイチジク属の固有種形成には、単なる地理的要因だけでなく、雑種形成やそれに伴う生物間相互作用の適応進化等の生態的要因が影響している可能性を示した。
おわりに
イチジク属とイチジクコバチとの共進化と共種分化のメカニズムを解明するためには、集団内の遺伝的多様性と分化および近縁種間の関係の解明がキーポイントの1つである。本研究では、イヌビワは地理的距離に伴った集団構造があることが示唆されたが、それが唯一の送粉者である送粉コバチの集団構造とは必ずしも一致しなかった。例えば、イヌビワでは関西と福岡、鹿児島と集団では分化の程度が低いのに対して、送粉コバチでは関西集団が固有ハプロタイプをもち、他集団との分化の程度は非常に高くなっていた。一方で、台湾と八重山集団の間では、短い距離であるにも関わらずイヌビワ、コバチ類共に高い分化があり、特に、コバチでは移入、移出が長期間断たれている可能性が示された。今後は、データから得られる様々な遺伝的分化のパターンが、地理的要因、生態的要因のどのようなバランスによって生じているのか、シミュレーション等を用いながら検討したい。