
地球上には数千万種の生物が存在していると推測されています。これほど多種多様な生物ですが、その起源を辿ってみるとただ1つの共通祖先から進化してきたものであることが分かります。我々は生物の系統進化(時間軸)と相互作用(空間軸)からその進化・多様化のプロセスとメカニズムを探っています。
最新の研究成果を含めたイチジク属植物とイチジクコバチのかけひきの妙が、季刊「生命誌」に掲載されています
107号 REAEARCH「持ちつ持たれつの生存戦略」
スライドや動画を使って詳しく説明しています
「イチジク属植物とイチジクコバチの共生関係の仕組みについて」
再び、オサムシ研究に取り組みはじめました
「オサムシ研究のこれまでとこれから」
NHK高校講座 基礎生物の取材を受けました
「第16回 遺伝子と私たち」遺伝子と形質の関係を具体的な例を用いてわかりやすく説明されています。
研究テーマ
イチジク属植物と昆虫類の相互作用と種分化機構
昆虫類は地球上で最も多様化した動物群です。動物の種数の約70%を占め、地球は昆虫の惑星であるとまで言われています。昆虫の種多様性をもたらしてきた様々な要因の中でも、植物との相互作用が最重要なものの一つです。特に昆虫と被子植物の相互作用は互いの種多様化に大きく寄与してきたと考えられています。多くの昆虫は花粉運搬と餌資源の獲得を通して被子植物と共生関係を構築しています。その顕著な例はイチジク属植物とイチジクコバチの共生系です。 イチジク属植物とイチジクコバチの共生関係には二つの特徴があります。一つは「絶対共生系」、つまり相手がいないと子孫を残すことができない、相互依存の繁殖システムのことです。イチジク植物はほぼ密閉状態の花嚢の内側に花を咲かせており、その花嚢を出入りできるイチジクコバチは唯一の花粉運搬者です。一方、コバチは一部の花に産卵し次世代の生息場と餌資源を得ることができます(図1)。もう一つの特徴は「高い種特異性」のことで、多くは「1種対1種」、つまり1種のイチジク植物に1種のコバチのみが送粉していることです。また、イチジクの花嚢には、送粉コバチの他に、寄生コバチや他の昆虫類なども棲息しているため、イチジク属植物を中心に非常に複雑な生態系が形成されています。 我々はこのイチジク属植物―イチジクコバチ共生系をモデル系として、生物の相互作用による種多様化のプロセスとメカニズムの解明を探っています。これまでに分子系統解析と集団遺伝解析、化学生態、形態分類などを通して、両者の共進化・共種分化の検証とそのプロセスの解明を行ってきたが、さらにゲノム解析や遺伝子発現解析の手法も加え、共生関係の構築・維持機構および共進化・種分化機構の分子メカニズムの解明を目指しています。また、寄主植物だけではなく非寄主植物に対する昆虫の識別機構についても研究を行っています。
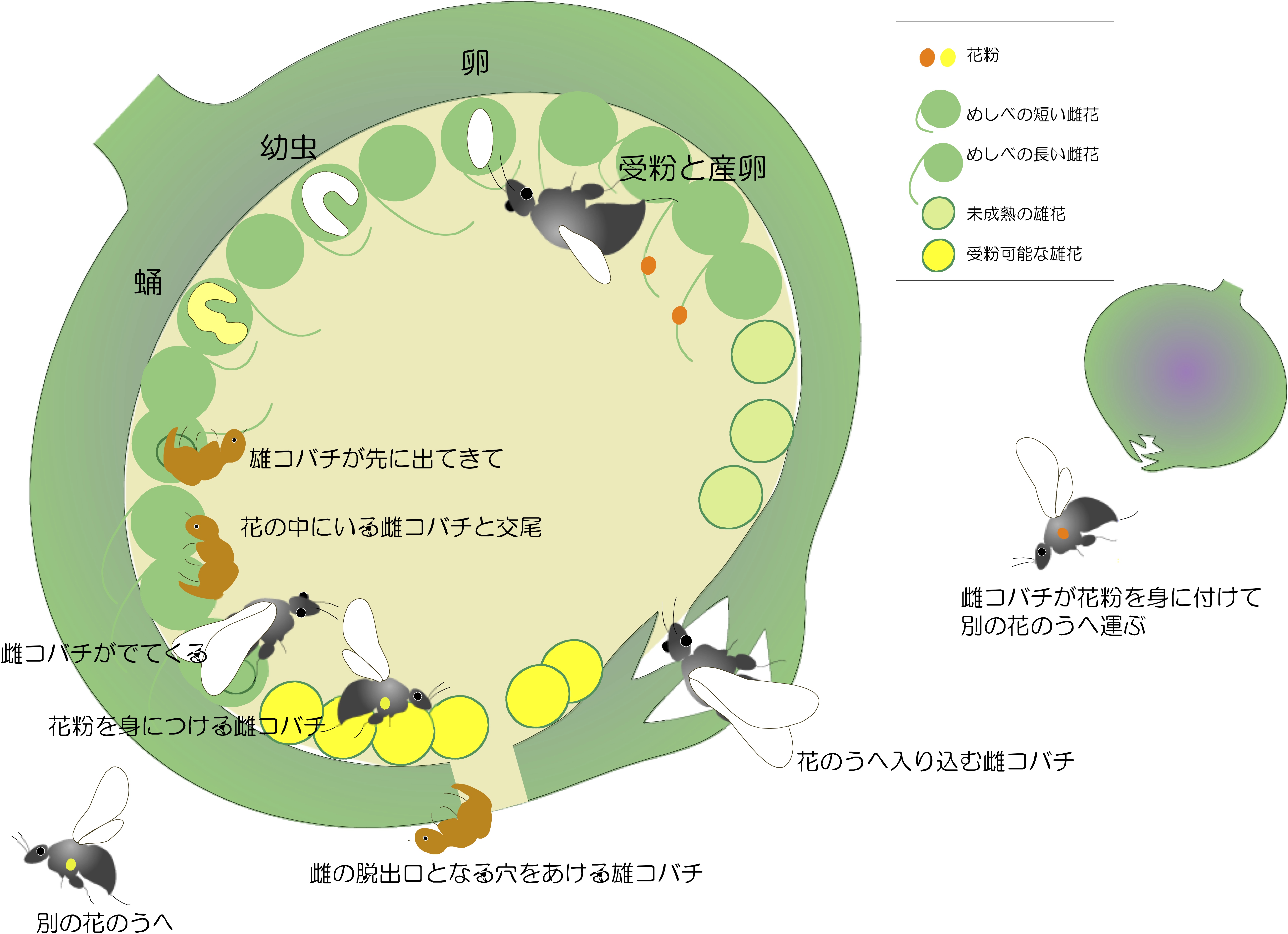
イチジク属植物とイチジクコバチの共生関係の仕組み
オサムシの後翅退化の分子機構の進化
生物の進化を理解するために、最も必要な情報は生物の系統関係です。系統樹の枝先に生物の形態や生態などの情報をマッピングすると、進化の様子が見えてきます。我々は1994年からおよそ10数年かけて、オサムシ(亜科)の分子系統解析を世界規模で行っていた。その結果から、オサムシの分布圏の成立や、地誌との関連、形態進化の様式などにおいて多くの新知見を得ることができました。しかし、オサムシの最大な特徴の一つである「飛べない」ことに関する進化的解明は、当時の研究技術の限りのためにできませんでした。 昆虫の進化・多様化の過程において翅の獲得は最重要なイベントです。昆虫と植物の相互作用が昆虫の多様化をもたらしてきた最大な要因の一つであると述べましたが、昆虫自身による翅の獲得も昆虫の多様化に大きく寄与したと考えられています。一方、現生の昆虫類では、甲虫類をはじめ、翅をなくして飛翔力を失ったものも少なくありません。また、翅の退化による飛翔力の喪失も種の多様性を促進していると考えられています。つまり、翅の獲得と退化はどちらも昆虫の種多様性に寄与している、実に興味深い現象です。オサムシ(亜科)は後翅を退化して飛べなくなった代表的昆虫群です(図2)。我々は昆虫類で広く見られる翅退化の進化的現象を理解するために、現在オサムシの後翅退化の分子メカニズムの解明を目指しています。

アオオサムシとその後翅の退化
Imura et al., 2018より改変
六脚類を中心とする節足動物の系統進化
生物の進化を理解するために、系統関係が最重要な情報であることは上でも述べましたが、もしその系統関係が間違っていたら、当然間違った進化の物語が導かれてしまいます。節足動物は、鋏角類(サソリ・クモなど)、多足類(ムカデ・ヤスデなど)、甲殻類(エビ・カニなど)と六脚類(昆虫類)に分類されています。節足動物に対する進化の理解は、近年の分子系統学の研究によって大きく変わりました。従来、六脚類は多足類に近縁で、陸上で誕生し進化してきたと考えられていたが、様々な分子情報を用いた系統解析の結果、六脚類は甲殻類に近縁であることが明らかになり、昆虫の共通祖先は陸上で誕生したのではなく、甲殻類の系統から分岐し、自ら陸上進出を果たしたことが分かりました。しかし、六脚類をはじめとする節足動物の系統関係についてはまだ不明なところが多く、それが節足動物の進化と多様化への理解の妨げになっています。 我々はこれまでに複数の核タンパク遺伝子を用いて、六脚類と多足類の目レベルの系統関係の解明を行ってきました(図3)。六脚類については、これまで提唱されていた内顎類、無尾類、無眼類の単系統性を否定し、分子系統学的にカマアシムシ目が六脚類において最も先に分岐した系統であることを示しました。また有翅昆虫の系統関係について、多新翅類の単一起源を強く支持する系統仮説を分子情報に基づいて初めて提唱し、完全変態類昆虫の目間の系統関係をほぼ完全に解明することができました。多足類については、ムカデ綱、ヤスデ綱、コムカデ綱 、エダヒゲムシ綱のうち、コムカデ綱が最初に分岐した系統であることを明らかにし、その分岐はこれまで考えたより遙かに古く、カンブリア紀初期に遡ることが判明しました。さらに、多足類の祖先種は、半増節変態を行っていたことと、体節と脚の数が少なかったことを推測することもできました。現在、より多くの分子情報によるゲノム系統解析の手法を用いて、節足動物の系統関係における未解決な問題の解明に取り組んでいます。
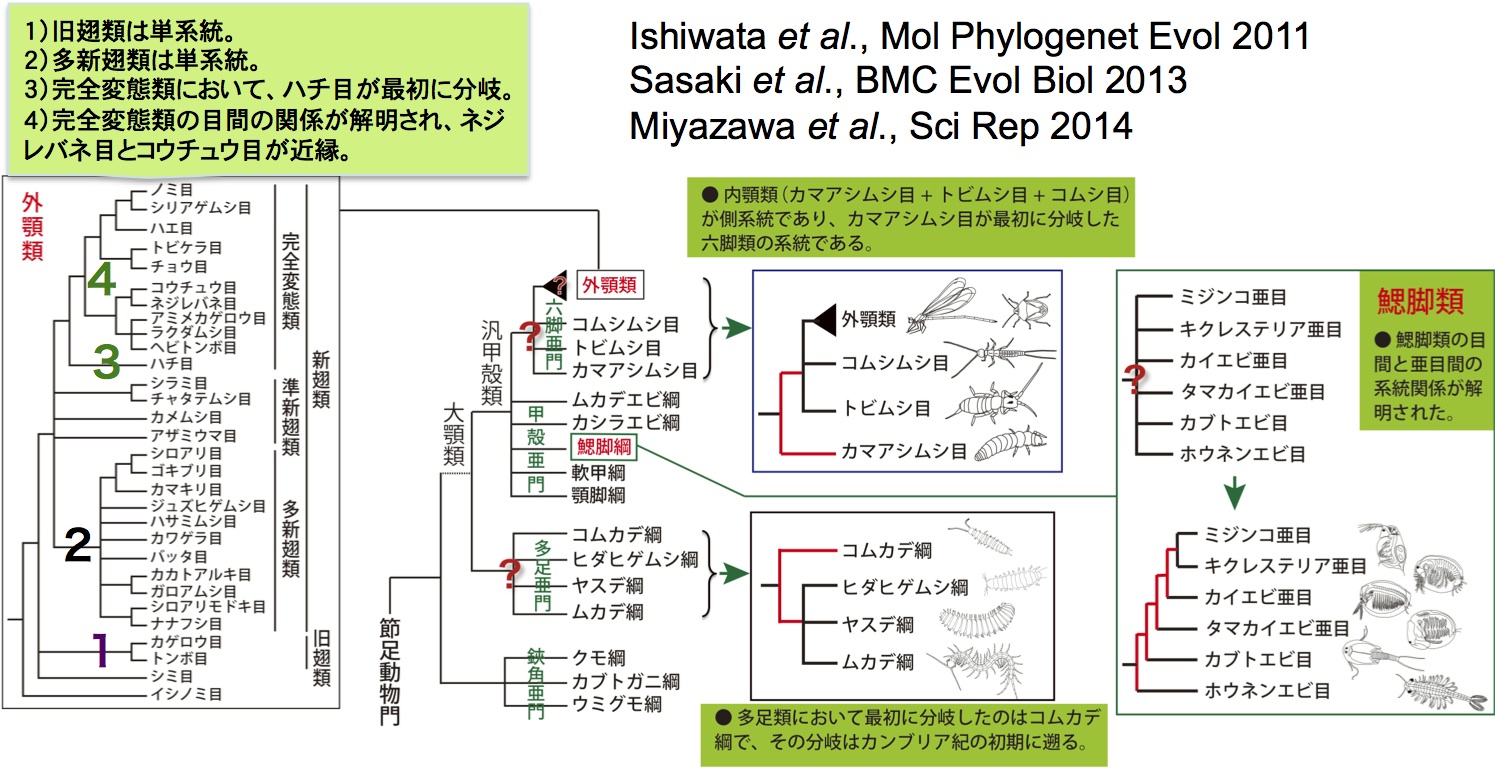
節足動物の系統進化
[最新論文] 論文一覧はこちら
Su Z.-H., Sasaki, A. Kusumi, J. Chou P.-A., Tzeng H.-Y., Li H.-Q., and Yu H. (2022)
Pollinator sharing, copollination, and speciation by host shifting among six closely related dioecious fig species
https://www.nature.com/articles/s42003-022-03223-0
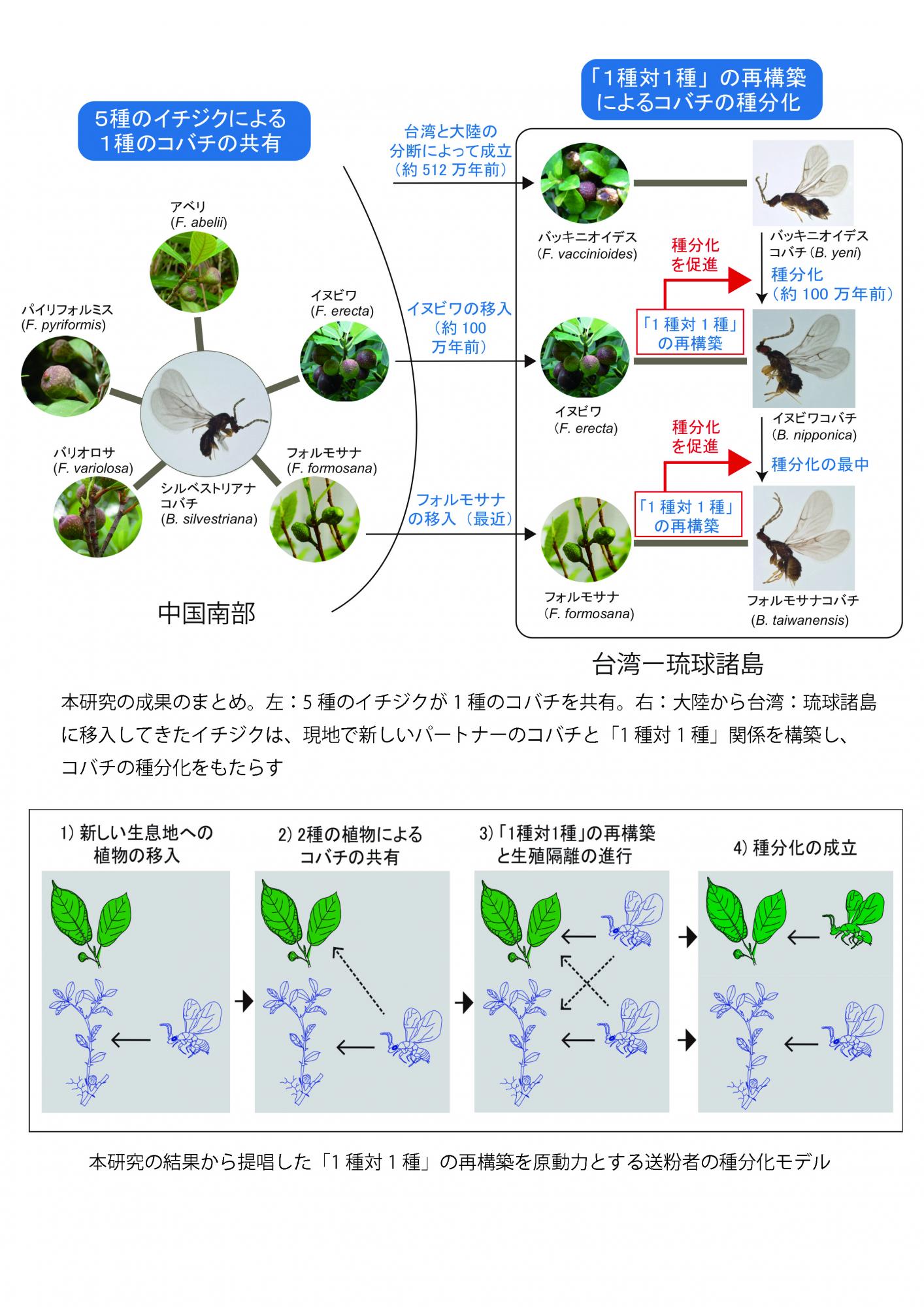
イチジク属植物(以下イチジク)とイチジクコバチ(以下コバチ)は、植物と昆虫が繁殖を相互に依存する絶対送粉共生系であり、両者が同調的に進化する「共進 化と共種分化」の典型的な例として古くから研究されてきました。しかし、 近年の研究からコバチとそのホストであるイチジクの種分岐が一致しない例が発見 され、コバチが宿主を変更するホストシフトによる種分化の可能性が示され ました。本研究は、コバチの種分化のメカニズムを明らかにするために、ホストシ フトが生じる可能性の高い島嶼地域に注目して、中国南部から台湾・琉球諸島にか けて分布する近縁のイチジク6種とそのコバチを研究対象として、それらの系統関 係と島嶼間の集団遺伝構造を様々な分子マーカーを用いて解析しました。そ の結果、1)中国南部の5種のイチジクの送粉者が 1 種のコバチであること、2) 大陸と島嶼でイチジクとコバチの移動によるホストシフトが生じた結果、同所的と異所的共送粉が起きたこと、3)大陸のイチジクが台湾・琉球諸島に移入後、現地 のコバチがホストシフトすることで新しい送粉者となり「1 種対 1 種」関係を再構 築した結果、コバチの種分化が生じていたこと、が判明しました。これらの研究結 果に基づいて、「1 種対 1 種」関係の再構築を原動力とする種分化モデルを提唱しました。
Okamoto, T. and Su, Z.-H. (2021)
Chemical analysis of floral scents in sympatric Ficus species: highlighting different compositions of floral scents in morphologically and phylogenetically close species.
https://doi.org/10.1007/s00606-021-01767-y
イチジク属植物の花の匂いの化学解析
イチジク属植物は、寄主特有のイチジクコバチによって受粉される。ほとんどの場合、1種のコバチが1種のイチジクの花粉を運んでおり、この高い種特異性はイチジクが放出する特定の花の匂いに対するコバチの応答によって維持される。我々は、同所的に生育する近縁な植物種間では、形態的な特徴が異なる種間よりも、形態的に似た種間で花の匂いに大きな違いが見られると仮説をたて検証を行った。仮説の検証のため、同所的に生育する5種のイチジクの花の匂いを捕集、ガスクロマトグラフ質量分析計で分析を行い、花の匂いの種間の違いを調べた。また、Y字管を用いてイチジクコバチの花の匂いに対する選好性を分析した。さらに、イチジクおよびイチジクコバチの種間の遺伝的距離、イチジクの形態的特徴の種間差を算出し、花の匂いの種間差との比較を行った。その結果、樹高やイチジクコバチの侵入口(花嚢の先端にある小さな穴)のサイズが類似しているイチジク種では、系統的に近縁であっても、明確に異なる花の匂いを放出していることが明らかになった。
.jpg)
Uozumi, T., Ishiwata, K., Grygier, M. J., Sanoamuang, L.-o. and Su Z.-H. (2021)
Three nuclear protein-coding genes corroborate a recent phylogenomic model of the Branchiopoda (Crustacea) and provide estimates of the divergence times of the major branchiopodan taxa
https://www.jstage.jst.go.jp/article/ggs/advpub/0/advpub_20-00046/_pdf/-char/en
3つの核蛋白遺伝子による鰓脚類の系統解析と分岐年代推定
鰓脚綱(甲殻亜門)は、6つの高次分類群、つまりホウネンエビ類、カブトエビ類、タマカイエビ類、カイエビ類、キクレステリアとミジンコ類によって構成され、形態とライフサイクルには多様性が富む。しかし、それらの分類群間の系統関係が長い間論争が続いていた。 本研究は、DNA合成酵素とRNA合成酵素をコードする3つのオーソロガス遺伝子のアミノ酸配列を用いて、鰓脚類の高次分類群の系統解析と分岐年代の推定を行った。その結果、鰓脚類の単系統性が強く支持され、また、高次分類群間の系統関係も明らかになった。まず、ホウネンエビ類が分岐し、次いで、カブトエビ類、タマカイエビ類、カイエビ類、キクレステリアとミジンコ類の順に分かれた。また、同じ遺伝子の塩基配列を用いた解析によって、ホウネンエビ類、ブトエビ類とミジンコ類の内部の種間の系統関係も明らかになった。さらに、アミノ酸配列データに基づく統計的検定を行い、これまでに提唱されていたカブトエビ類の系統的位置のいくつかの仮説を却下した。一方、本研究の結果はトランスクリプトームによる最近の研究結果とも一致しており、本研究に用いた3つの遺伝子による鰓脚類の高次分類群の系統解析の有効性が確認された。化石データに基づく分岐年代推定では、現生の鰓脚類の最初の分岐がカンブリア紀初期に遡ることが判明した。また、主要な分類群が分岐した後、それらの分類群内の多様化が長期間に渡り進んでおらず、ペルム紀後期以降に始まったことが示唆された。

上:本研究で明らかにした鰓脚類の系統関係と分岐年代
下:本研究に用いた鰓脚類各種
New data on lance flies (Diptera, Lonchaeidae) associated with figs (Moraceae, Ficus spp.) in Japan and Taiwan, with descriptions of two new species of the genus Silba Macquar
J. Asia Pac. Entomol. 23 : 364-370.

Evolutionary history of carabid ground beetles with special reference to morphological variations of the hind-wings.
Proc. Jpn. Acad., Ser. B 94: 360-371.
 日本産オサムシの後翅の退化
日本産オサムシの後翅の退化Wachi, N., Kusumi, J., Tzeng, H.-Y., and Su Z.-H. (2016)
Genome-wide sequence data suggest the possibility of pollinator sharing by host shift in dioecious figs (Moraceae, Ficus).
Mol. Ecol. 25: 5732-5746. doi:10.1111/mec.13876.
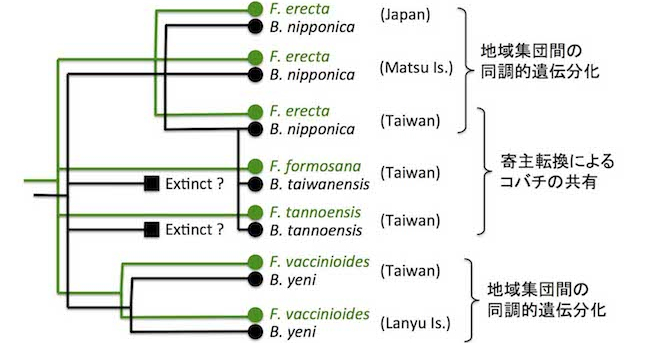 イヌビワ Ficus erecta とその近縁種(緑色)およびそれらの送粉コバチ(黒色)の系統関係
イヌビワ Ficus erecta とその近縁種(緑色)およびそれらの送粉コバチ(黒色)の系統関係
Osawa, S., Su Z.-H. Nishikawa, M., and Tominaga, O. (2016)
Silent evolution.
Proc. Jpn. Acad., Ser. B 2: 455-461. doi:10.2183/pjab.92.455.
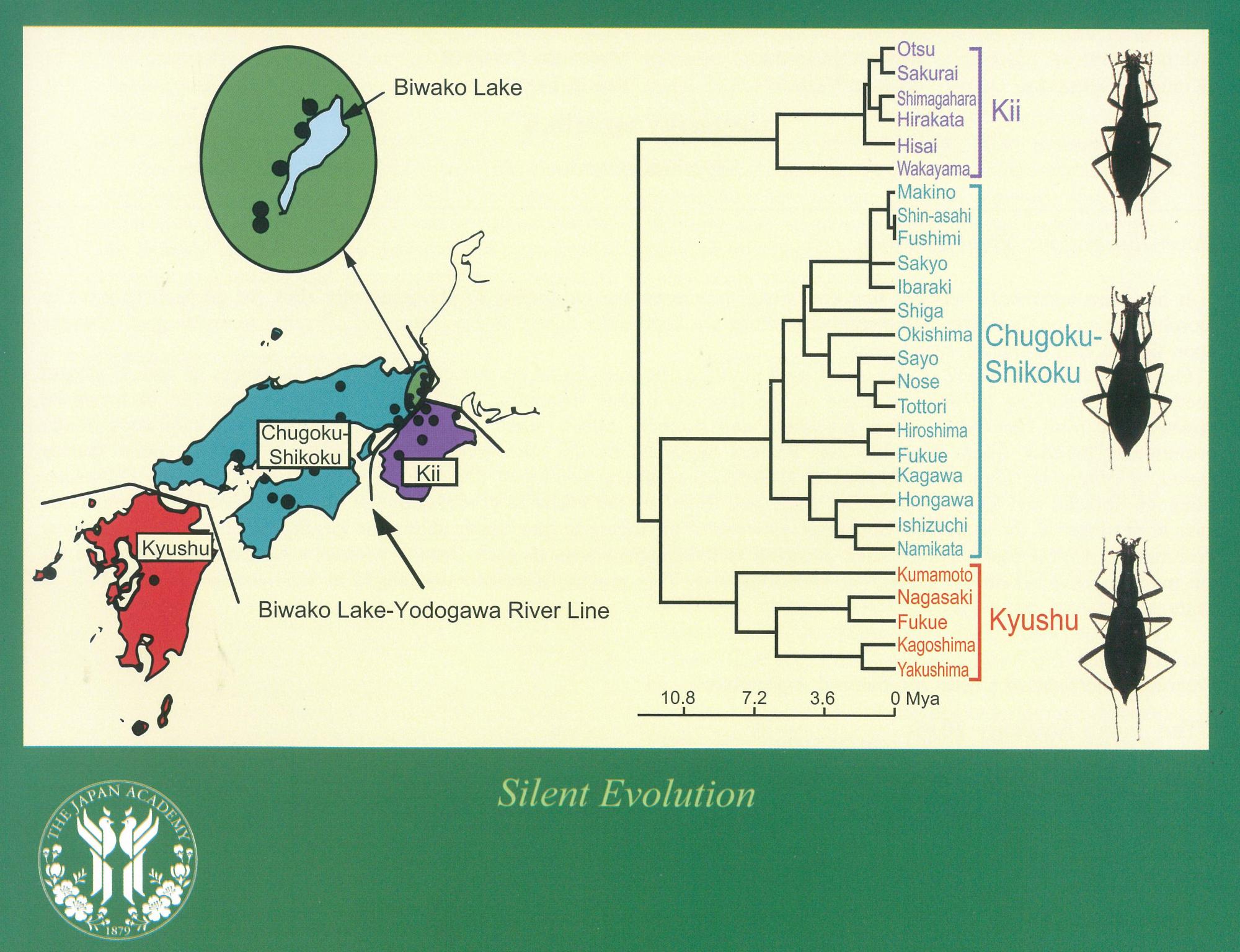
ホンマイマイカブリ Damaster blaptoides blaptoides の3つの地域系統
年度別活動報告一覧はこちら
ラボ日記