年度別活動報告
年度別活動報告書:2015年度
分子系統から生物進化を探る 3-1.昆虫と植物との共生・共進化・種分化機構の解明
蘇 智慧(主任研究員) 和智仲是(奨励研究員)
佐々木綾子(研究補助員) 南紘彰(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士あるいは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は互いに相互適応的な関係を築くことによって促されてきたように見える。その最たる例が「1種対1種」の密接な送粉共生関係を築くイチジク属植物 (クワ科) と送粉コバチ(イチジクコバチ科、以下イチジクコバチ)であり、それぞれ数百種が知られている。我々は現在このイチジク属植物とイチジクコバチとの共生関係の構築過程と維持機構およびその共生系における種分化機構などについて研究を行っている。
イチジク属植物は、東南アジア・アフリカ・中南米など熱帯を中心に分布している。日本はイチジク属植物の分布域の北限にあたり、南西諸島を中心に16種が分布している。イチジク属植物は花嚢とよばれる袋状の閉じた花序をつけ、その内側に多数の花を咲かせる。送粉者であるイチジクコバチは、この花嚢に入り花粉を媒介するだけでなく同時に花に産卵する。ふ化した幼虫は虫こぶ(昆虫が改変した植物の組織)の中で成長する。やがて次世代のイチジクコバチが花嚢内で羽化・交配し、雌成虫が花粉を持って他の花嚢へと移動することで次の受粉が成立する。このようにイチジク属植物とイチジクコバチの二者は、繁殖において互いに強く依存し合った関係と言える。
イチジク属植物とイチジクコバチとの共生関係は、「1種対1種」という種特異性が極めて高いものと言われている。この「1種対1種」関係を維持しながら種分化が起きるとしたら、同調した種分化や系統分化が起こることが予想される。これまで分子系統学的解析を用いてこの仮説を検証する研究が行われてきた。その結果、イチジク属植物の節(植物では属・亜属・節・種と分類する)レベルの系統関係とイチジクコバチの属レベルの系統関係がおおまかに一致し、仮説が支持されている。しかし種間・種内レベルでの系統関係の矛盾のほかに、イチジクコバチの隠蔽種や1種のイチジク属植物に複数種のイチジクコバチ(あるいはその逆)が共生するなど、不明瞭な点が残っている1-4)。我々のこれまでの研究によって、日本産のイチジク属植物とイチジクコバチでは「1種対1種」関係の厳密性が見られ、同調的系統分化が示唆された5)。一方でメキシコ産のイチジク属植物とイチジクコバチでは、近縁種間で「1種対1種」関係の乱れが見られた2)。また、小笠原諸島産のイチジク属植物の進化・種分化の過程においては異種間交雑が起きていたことが示唆され、雑種形成がイチジク属植物の種分化をもたらす要因の1つであると考えられた6)。また、イチジク属植物が特定のイチジクコバチを自らの花へ呼び寄せるための手段として、花の匂い(花から放出される分子量300以下の揮発性に富んだ化学物質の集まり)が注目されてきた。多くの送粉者は匂いを嗅覚によって捉え花の探索に利用している。イチジク属植物でも、花嚢の外見は緑色で非常に目立ちにくいため視覚情報よりも嗅覚情報としての花の匂いが有効であると考えられる。
このような背景のもと、昨年度に引き続き、本年度も主にイヌビワとその近縁種およびそれぞれのイチジクコバチの種間・種内の関係に注目して、集団遺伝学的解析、花の匂いを認識する遺伝子の探索を行った。
結果と考察
1. イヌビワコバチとその近縁種の寄主植物への適応を決定づけるゲノム領域の特定
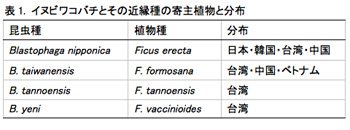
イヌビワ(Ficus erecta)は東南アジアから東アジアにかけて広く分布しており、日本・韓国・台湾・中国から知られている。さらに台湾にはイヌビワに近縁な3種 F. formosana, F. tannoensis とF. vaccinioides が分布している。これら4種の近縁な植物種に対して、それぞれ種特異的に共生関係にあるイチジクコバチが知られている(表1)。植物種は形態的・遺伝的に明確に区別ができる。その一方で、F. vaccinioides を利用する Blastophaga yeni を除くイチジクコバチ3種(B. nipponica, B. taiwanensis, B. tannoensis)は、形態的によく似ており、記載時に用いられた種の識別形質も連続的で曖昧である。これまで、イチジクの種間関係・コバチの種間関係について詳細な解析を行ってきた。
これまでの研究により、下記のことが明らかになっている。
1) イチジクは、遺伝的にも花の匂い物質の組成も分化している(2011, 2012, 2014年度活動報告)
2) Blastophaga nipponicaに地域的分化が見られる(2013, 2014年度活動報告)
3) 限られた地域に分布するBlastophaga yeniに分集団化が見られる(2013, 2014年度活動報告)
4) 台湾に分布する3種のコバチ(B. nipponica台湾集団, B. taiwanensis, B. tannoensis)は、イチジクに対応した分化が見られない(2013, 2014年度活動報告)
本年度は寄主植物への適応に関連した領域を探すため、種間で特に大きな分化を示すゲノム領域の探索を行った。寄主植物が異なるにもかかわらず、遺伝的分化がほとんどない近縁種間で見出される遺伝的分化の大きなゲノム領域は、寄主植物の違いを決定づけている領域である可能性が高い。明確な遺伝的分化が見られない台湾のイヌビワコバチB. nipponicaとその近縁種(B. taiwanensis, B. tannoensis)間で遺伝的分化が大きい領域は合計46領域あった。これらのゲノム領域周辺に寄主植物への適応を決定づける領域があると考え、候補領域とした。これまで決定したドラフトゲノムに対して、相同性検索を行い、これら46候補領域のゲノム上の位置を明らかにしようと試みた。その結果、37領域と相同な領域を含む102コンティグ配列(ゲノム上のひとつながりの塩基配列)が見出された。さらにそのうち、遺伝子と考えられる領域を含むコンティグ配列は48であった。この48コンティグ配列のうち、1つの候補領域が1つの遺伝子を含むコンティグ配列上にのみに位置すると考えられたものは12領域であった。これら12コンティグ配列の中で候補領域は、10ヵ所は遺伝子間領域、2ヵ所は遺伝子内のイントロンに位置していた。さらにこの12コンティグ配列のうち、9つが属もしくは種特異的に発現している遺伝子も含んでいた。
コンティグ配列に含まれる遺伝子の機能を予測したところ、2つのコンティグ配列がホメオボックス遺伝子を含むと推測された。このうちホメオボックス遺伝子 caupolican に相同性がある配列の近くに位置する候補領域は、どの種間でも遺伝的分化が大きい傾向にあった。ショウジョウバエではホメオボックス遺伝子の一つであるacj6の変異体は嗅覚受容に異常が見られ、またacj6は嗅覚受容体の発現に影響している7)。さらにカイコにおいてacj6と相同な遺伝子の変異体は嗅覚受容やフェロモン受容に異常が見られる8)。イチジクコバチも同様にホメオボックス遺伝子に関連した変異が、嗅覚受容に影響し寄主植物への適応を決定づけているのかもしれない。
また、Low-density lipoprotein receptor-related protein 2に相同性がある配列の近くに位置する候補領域は種ごとに遺伝的分化が大きいわけではなかったが、種特異的な変異をもち胸腹部で発現している遺伝子の一つだった。転写調節だけでなく遺伝子自体の違いも寄主植物の違いに影響を与えていることを示唆しているのかもしれない。
今後、イヌビワコバチのゲノム情報を充実させることで、今回ゲノム上の位置が特定できなかった9領域を含めた、それぞれの候補領域の位置関係が明らかになると期待している。今回検出された近傍に遺伝子を含む12領域については、サンプル数を増やし、さらに詳細に種間の遺伝的分化の程度を調べる予定である。
2. イヌビワコバチのゲノム解読
イヌビワコバチの研究を効率的に進める上で、ゲノムに関する情報の取得と解析は欠かすことができない。近縁なイチジクコバチ(Ceratosolen solmsi)では全ゲノムが解読されている9)が、公開が限定的であるため配列情報を自由に使える状況でない。
そこで、より効率的に研究を展開するために、次世代シーケンサ(Ilumina, Miseq)を用い、イヌビワコバチのゲノム解読を試みている。これまでに得られた、無作為に断片化したゲノムDNAから構築したライブラリー(Nextera)を用いたMiseq 2ラン分の情報にさらに、ddRAD-seqの配列情報を加えた。これらの配列のアセンブル(相同な配列をまとめること)を行った結果、コンティグ配列が46,359個(最大で31.5万塩基)得られた。今後、さらに異なる断片長(1Kb, 2Kb, 5kbなど)のゲノムライブラリを作成し、より網羅性の高い充実したゲノム情報にできればと考えている。さらに、ゲノム比較を行うために、イヌビワコバチの近縁種(B. taiwanensis, B. tannoensis, B. yeni, Blastophaga sp. 1, Blastophaga sp. 2, Blastophaga sp. 3)についてドラフトゲノム情報も合わせて取得する予定である。
3. イヌビワコバチとその近縁種の寄主植物認識遺伝子の探索
イチジクコバチはそれぞれ特異的なイチジク属植物を利用するために形態的にも生態的にも適応しているように見える。そして、寄主植物種を認識するのには嗅覚情報を利用しているのではないかと考えられている。しかし、イチジクコバチにおいて、寄主植物認識にどのような遺伝子が重要な役割を担っているかという具体的な機構は十分には明らかになっていない。この寄主植物認識遺伝子を明らかにすることができれば、イチジクコバチがどのようにして寄主植物を認識し、どうやって適応してきたかという共生の進化についての重要な知見が得られることが期待される。
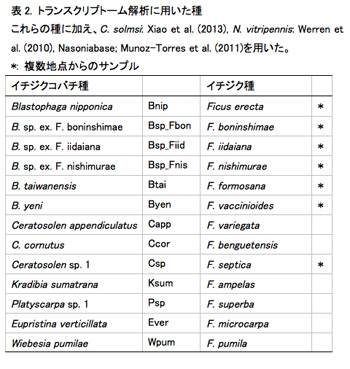
イヌビワコバチとその近縁種を含む13種(表2)の頭部、胸腹部で発現している遺伝子群を網羅的に比較することにより、主に嗅覚情報の認識に関与している可能性のある遺伝子の探索・同定を試みた。12種の雌の生体頭部、胸腹部からRNAを抽出し、cDNAを合成した。合成したcDNAをライブラリー化し、次世代シーケンサ(Miseq)で解析を行った。得られた配列データをゲノムが解読されているコバチ(Nasonia vitripennis, C. solmsi)の遺伝子配列と比較した。その結果、頭部・胸腹部のそれぞれで約1,800万配列を用いて約5,000の相同な遺伝子配列とその発現情報が得られた。
得られた情報を用いて種間で比較した結果、Blastophaga属・Ceratosolen属で属特異的に発現している遺伝子が頭部で合計56、胸腹部で合計45検出された。一方で B. nipponica, B. sp. 1 ex. F. boninsimaeで種特異的に発現している遺伝子は頭部で合計167・胸腹部で合計109検出された。また、Blastophaga属・Ceratosolen属で属特異的な変異を持つ遺伝子が頭部で40・胸腹部で53、B. nipponica, B. sp. 1 ex. F. boninsimaeで種特異的な変異を持つ遺伝子が頭部で1・胸腹部で3検出された。これらの遺伝子のゲノム上の位置と種間で遺伝的分化が大きい領域の位置を比較することで寄主植物への適応を決めるゲノム領域を検出できると期待される。
さらに、得られたトランスクリプトームの情報を用いて系統関係を推定する方法を確立した。イチジクコバチにおいて得られた配列を用いて系統関係を推定したところ、約800〜1000の遺伝子座の情報による支持率の高い系統関係が推定された。トランスクリプトーム情報の得られる種であれば、同様の方法を用いて、信頼性の高い系統関係を推定できることが期待できる。
4. イヌビワの雄株の花嚢に種子が作られる?
イチジク属植物には、雌雄同株と雌雄異株の種がある。雌雄同株の花嚢には雄花、花柱の長い雌花(種子生産)と花柱の短い雌花(コバチ育ち)が含まれる。一方、雌雄異株の種は、雄株の花嚢に雄花と花柱の短い雌花が、雌株の花嚢に花柱の長い雌花のみが存在する。つまり、雌雄異株の種では、種子生産とコバチ育ちが別々の樹に分かれて分業という形になっている。
分子系統解析の結果から、イチジク属植物の祖先は雌雄同株であり、進化の過程で雌雄異株の種が出現し、また少なくとも一回は雌雄異株から雌雄同株に逆戻りの進化が起きていると考えられている。
このような「雌雄同株—雌雄異株」の間の進化はどのようにして起きるのだろうか。そのメカニズムは全く分かっていない。BRH食草園に育てているイヌビワ (F. erecta) の観察から興味深い現象が発見された。イヌビワは雌雄異株の種であるため、雄株の花嚢にはコバチが育つ、花柱の短い雌花と雄花だけがつくことになっているはずである。しかし、その雄株の熟した花嚢から種子が見つかり、その種子が発芽して実生になった(図5)。この発見から2つの可能性が考えられる。1つはコバチを育てる、花柱の短い雌花でも産卵されなければ、花粉がつくと種子が作られる可能性である。しかし、もしこの可能性が本当であれば、このような現象はもっと多く観察されてもおかしくないが、そのような報告は知られていない。また、イヌビワの花柱の長い雌花と花柱の短い雌花の形態を比較してみると、花柱の長さの違いの他に、花柱の先端の形状も異なっているため、2種類の雌花はすでに機能分化しているかもしれない。もう一つはその雄株の花嚢に種子を作る雌花が混じっている可能性である。もしこれは何かの遺伝的突然変異によって生じた現象であれば、まさに雌雄異株から雌雄同株への進化に関わる遺伝的変異であり、「雌雄同株—雌雄異株」の間の進化の分子メカニズムの解明に繋がる。今後、イヌビワの花嚢内の花の形態の観察を含め、この現象を更なる追求し、そのメカニズムの解明も探っていきたい。
おわりに
イチジク属植物とイチジクコバチとの共生・共種分化機構を解明するために、これまで主として分子系統学・集団遺伝学・化学生態学の手法を用いて研究を行ってきた。昨年度に引き続き、本年度も主にイヌビワとその近縁種およびそれぞれのイチジクコバチの種間・種内の関係に注目して、集団遺伝学的解析を行った。予備的ではあるが、近縁種間で遺伝的分化の大きな領域と、複数種のトランスクリプトーム情報を比較する方法を用いた解析により得られた種特異的な発現を示す遺伝子の両方が位置するゲノム領域12カ所を特定した。それらの領域の近傍には、転写因子を含むものがあった。転写調節の違いが寄主特異性を制限している可能性を示唆しているのではないかと考えている。今後、これらの領域について、サンプル数を増やしても種間で遺伝的分化が見られるか追試を行っていく予定である。今後の目標としてイヌビワコバチの全ゲノム情報の解読・イヌビワコバチの近縁種についてドラフトゲノム情報の取得が挙げられる。今後、新たに得られるゲノム情報とこれまでの結果を統合し、ゲノム上のいくつの、どのような領域が異なることでイチジクコバチが特定の寄主植物に適応しているかを明らかにすることを目指したい。
また、今回、イヌビワの雄株の花嚢に種子が作られる興味深い現象を発見した。このことは雌雄異株から雌雄同株への進化に関わる遺伝的変異が生じた可能性を示しており、今後さらに詳細に調べていく予定である。
一連の解析には下記を用いた。
配列データ解析:perlスクリプト、pythonスクリプト; ddRAD-seqのデータ解析:Stacks, pyRAD; 集団構造の推定:STRUCTURE; 系統関係の推定:RaxML; 次世代データの解析: CLC Genomics Workbench; 集団間分化の推定: Popgenome; 相同性検索: blastn, tblastx; Gene Ontology検索: blast2go; 遺伝子モデル予測:AUGUSTUS