年度別活動報告
年度別活動報告書:2012年度
分子系統から生物進化を探る 3-1.イチジク属植物とイチジクコバチとの共生・共進化
蘇 智慧(主任研究員) 岡本朋子(奨励研究員)
佐々木綾子(研究補助員) 宮澤秀幸(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。その最たる例が「1種対1種」の密接な送粉共生関係を築くイチジク属植物 (クワ科) とイチジクコバチ科昆虫であり、双方が800種以上の種数を誇っている。我々は現在イチジク属植物とイチジクコバチとの共生関係の構築と維持のメカニズム、およびその共生系における種分化様式などについて研究を行っている。
イチジク属植物は、東南アジア、アフリカ、中南米など熱帯を中心に分布している。日本はイチジク属の分布域の北限にあたり、南西諸島を中心に16種が生息している。イチジク属植物は花嚢とよばれる袋状の閉じた花序をつけ、その内側に多数の花を咲かせる。送粉者であるイチジクコバチは、花嚢にはいった場合、授粉と同時に花に産卵し、ふ化した幼虫は子房を食べて成長する。やがて次世代のコバチが花嚢内で羽化・交配し、雌成虫が花粉を持って他の花嚢へと移動することで受粉が成立する。このようにイチジクとイチジクコバチの2者は、繁殖を互いに強く依存し合った関係といえる。
イチジク属植物と送粉コバチとの共生関係は、「1種対1種」という種特異性が極めて高いものと言われている。この「1種対1種」関係を維持しながら種分化が起きるとしたら、同調した種分化や系統分化が起こることが予想される。これまで分子系統学的解析を用いてこの仮説を検証する研究が行われてきた。その結果、イチジク属の節(section)レベルの系統関係とコバチ類の属のそれとがおおまかに一致し、仮説が支持されるものの、種間、種内レベルでは系統関係の矛盾のほかに、送粉コバチの隠蔽種や1種のイチジク属植物に複数種の送粉コバチ(或いはその逆)が共生するなど、不明瞭な点が残っている1-3)。我々の研究において、日本産のイチジク属植物とイチジクコバチでは、「1種対1種」関係の厳密性が見られ、同調的系統分化が示唆された4)が、メキシコ産の材料の解析では、近縁種間で「1種対1種」関係の乱れが示された2)。また、イチジク属植物の進化・種分化の過程において交雑が起きていたことも示唆され、雑種形成がイチジク属植物の種分化をもたらす要因の1つであると考えられる5)。また、イチジク属植物が特定のパートナーを花へ呼び寄せるために用いるシグナルとして、主に嗅覚情報である花の匂いが注目されてきた。花の匂いとは、花から放出された分子量300以下の揮発性に富んだ化学物質(の集まり)を指す。イチジクの花嚢の外見は緑色で非常に目立ちにくいため、視覚情報よりも嗅覚情報が有効であると考えられている。本年度は主にイヌビワとオオバイヌビワおよびそれらの送粉コバチの種内の集団間の関係を注目して、分子系統解析、集団遺伝解析、花の匂い物質の解析を通して研究を進めてきた。
結果と考察
1)イヌビワ(Ficus erecta)とイヌビワコバチ(Blastophaga nipponica) の種内レベルの遺伝的多様性、集団間の遺伝的分化の解析
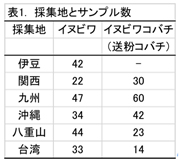
イヌビワは広い分布域をもち、日本、済州島(韓国)、台湾、中国南部とベトナム北部に生息している。本研究では、日本の5地域(伊豆、関西、九州、沖縄、八重山)と台湾で採集を行った。各植物個体の葉および花嚢を採集し、葉の一部を用いてDNAを抽出した。イヌビワコバチは、植物1個体あたり、1〜4個の花嚢から採取した。同じ親由来の個体が重複するのを避けるため、花嚢1つあたり1匹の雌を用いることとし、1個体ごとにDNAを抽出した。日本の集団に関しては、統計的解析に必要な数のサンプルを回収した(表1)。
1.1. 葉緑体DNAによるネットワーク解析とハプロタイプの分布
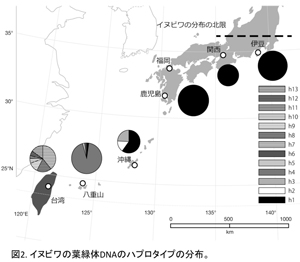
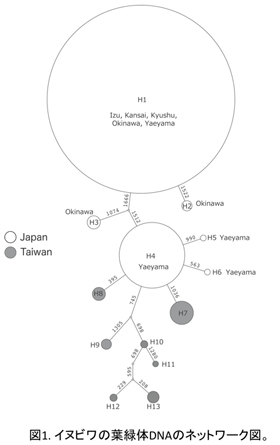
葉緑体DNA 6 領域(rps16イントロン、trnGイントロン、petBイントロン、trnLイントロン、trnL-trnFスペーサー、atpB-rbcLスペーサー)およそ3500塩基を用いて解析を行った。解析ソフトは TCS v.1.21である。その結果は図1に示し、各地域におけるハプロタイプの分布は図2にまとめた。イヌビワには合計13ハプロタイプが見つかった。日本産のイヌビワは6ハプロタイプ(H1-H6)を有しているが、そのうちの2つ(H1とH4)はメインである(八重山タイプと本州・九州・沖縄タイプ)。本州のイヌビワは九州から伊豆まで広い範囲にわたっているにもかかわらず、1つのハプロタイプ(H1)しか見つからない。沖縄の集団は3つのハプロタイプ(H1-H3)を有しているが、そのうちの一つ(H1、半分以上を占める)は本州・九州タイプと同じもので、他の二つは沖縄特有のものである。八重山集団には4種のハプロタイプ(H1, H4-H6))が見つかり、そのうちの一つ(H4)は八重山特有なもので、90%以上を占めている。ハプロタイプH1は日本産全集団が共通に有するものである。この結果から、日本産のイヌビワは南方から北進して分布域を拡大し、本州に到達したあと、比較的短期間で広がったと考えられる。一方、日本産イヌビワと比較して台湾産のものは、分布域が狭いにもかかわらず7つのハプロタイプ(H7-H13)が見つかった。日本産のイヌビワと共通に持つハプロタイプは見つからなかった。この結果は台湾産のイヌビワはより多様性に富み、日本産イヌビワとの間にある程度分化が起きていることを示唆している。
1.2. マイクロサテライトマーカーを用いたイヌビワの遺伝的構成の解析
イヌビワの塩基配列多型は非常に少ないことが示されていたため、集団間の分化の有無を解析するには、より多型性の高いマイクロサテライトマーカーを用いる必要があった。前年度までに9つのマイクロサテライトマーカーを用いて、関西、九州、沖縄、八重山と台湾のサンプルの解析を行った。本年度は新たにイヌビワの分布北限にたる関東(伊豆)の集団を加え、また台湾集団の個体数の追加をして解析を行った。
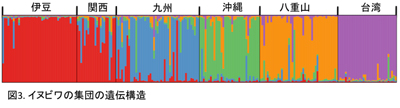
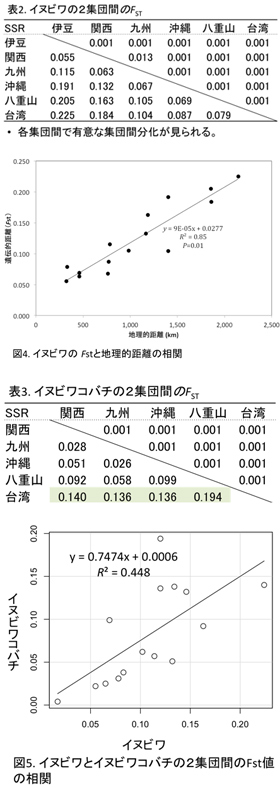
マイクロサテライトデータの解析は、Structure ver2.3.2、GeneAlEx6.3とArlequin ver 3.11というソフトウエアを用いて行った。イヌビワの遺伝構成を解析したところ、5つの遺伝的集団が確認され、それらは地域的集団と一致することが判明した。関西は伊豆と九州が混合したような集団になっており、独立的な遺伝的集団として判断するのは困難である。また、各集団において隣接した集団間での遺伝子流動の形跡が見られる(図3)。
また、集団間の遺伝的分化の程度を示す統計量(Fst)を計算したところ、地域集団間の遺伝的分化が有意にあることが示された(表2)。さらに、2つの地域集団間のFstとその地理的距離には有意な相関が見られた (Mantel test p<0.01)(図4)。従って、イヌビワの遺伝子流動(花粉や種の移動)は地理的距離が大きいほど制限される度合いが強いことが予想された。
一方、イヌビワコバチの集団間の遺伝的分化程度を示す統計量 (Fst) を解析したところ、イヌビワと同様に集団間の遺伝的分化が起きていることがわかった。特に台湾集団と日本の集団との間では、遺伝的分化が高い(表3)。しかし、Fstと地理的距離との相関はイヌビワと違って低い。
イヌビワとイヌビワコバチの集団構造、集団間分化の程度を比較すると、イヌビワの方がより地域集団に分化しやすい傾向があることが示唆された。イヌビワとイヌビワコバチの日本、台湾集団では遺伝的多様性の程度や2集団間のFstの値の相関関係を解析したところ、Fst の値に相関がみられることが判明した(図5)。この結果から、イヌビワとイヌビワコバチの地域集団間の遺伝子流動は相互に影響しあっている可能性が高いと考えられる。
2)オオバイヌビワ(Ficus septica)とオオバイヌビワコバチ(Ceratosolen bisulcatus) の種内の集団間の分化
イチジクの花粉を運ぶイチジクコバチは、花の匂いを用いて寄主を識別し、訪れることが知られている。すなわち、花の匂いはイチジクとイチジクコバチの両者が繁殖を達成するために重要なシグナルであるといえる。植物の遺伝子流動が限定された集団間では、それぞれの地域で匂いの特殊化が起こり、それが集団間の生殖隔離に寄与する可能性が考えられる。本研究では、オオバイヌビワとその送粉者であるオオバイヌビワコバチを対象とした集団遺伝学的解析によって、集団間の遺伝的交流の度合いを調べ、さらに遺伝的に分断される傾向にある集団間での花の匂いの比較を行った。
雌雄異株であるオオバイヌビワは、日本の奄美大島から台湾、中国南部、ベトナム、マレーシア、オーストラリアと広域にわたって分布することが知られている。日本から台湾の北部においては、オオバイヌビワコバチによって花粉が運搬されており、両者の間には極めて高い種特異性が見られる。オオバイヌビワおよびオオバイヌビワコバチの集団間における遺伝的交流の度合いを調べるため、マイクロサテライトを用いて、集団間の遺伝構造および遺伝子流動の程度について調べた。サンプルの採集は沖縄、八重山と台湾で行った(図6)。
さらに、オオバイヌビワの開花中の花嚢 (3から6個) をガラスバイアルに入れ、吸着剤(TENAX-TA)を用いたダイナミックヘッドスペース法によって花の匂いの捕集を行った6)。吸着された匂いは、熱脱離システムを用いてガスクロマトグラフ質量分析計(GC-MS)に導入し、定性的に分析を行った。GC-MS分析によって得られた匂い物質の組成比データから、サンプル間の(非)類似度(Bray-Curtis dissimilarity index)を算出し、それらを非計量多次元尺度構成法(Non-metric MultiDimensional Scaling)を用いて2次元平面上にプロットし、比較を行った。
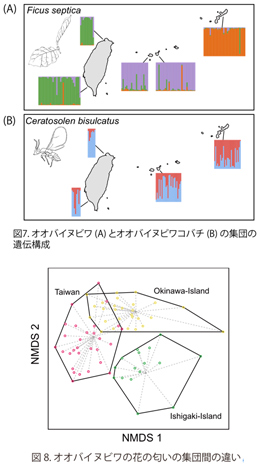
マイクロサテライト8遺伝子座を対象としたオオバイヌビワの集団遺伝学的解析の結果、台湾内では遺伝的な変異は観察されなかったものの、台湾–石垣島–沖縄島の間では遺伝構成が異なることが明らかになった(図7)。また同様に、花の匂いの分析の結果、オオバイヌビワの花の匂いは、台湾–石垣島–沖縄島の間では明瞭な違いが見られたが、台湾内では違いがみられなかった(図8)。本研究で調査を行った石垣島と台湾の間は約300kmであり、これは台湾の北部と南部の距離(約400km)とほとんど変わらない。それにも関わらず、台湾と石垣島の間ではオオバイヌビワは遺伝的分化を示し、さらに花の匂いにも違いがあることが明らかになった。台湾内では、オオバイヌビワは道沿いを中心に連続的に生育し、明瞭な分布の分断が見られない。それによって、集団間での遺伝的な交流が保たれている可能性が高いと考えられる。一方、海によって集団が分断された島間では、遺伝子の流動が制限されると考えられる。一方、オオバイヌビワの送粉者であるオオバイヌビワコバチのマイクロサテライト解析の結果、台湾と沖縄島の間で緩やかな遺伝的分化の傾向がみられたものの、島間における明瞭な遺伝構造の違いは見られなかった(図7)。イチジクとイチジクコバチの送粉共生系では、イチジクコバチは寄主のイチジクの花粉を持って分散し、その後自らも繁殖を行う。そのためイチジクとイチジクコバチの遺伝分散の距離は多くの場合一致すると考えられるものの、本研究の結果では明らかな不一致がみられた。これは、イチジクとイチジクコバチの間で、分布拡大のパターンが異なるなどの可能性が考えられる。今後調査を進めていくことで、イチジクとイチジクコバチの種分化のメカニズムの解明に繋がると期待される。
おわりに
イチジク属とイチジクコバチとの共進化と共種分化機構を解明するためには、これまで主として分子系統解析と集団遺伝学的解析の手法を用いて行ってきた。両者の間に、同調的な種分化が示される一方、一致しない結果も見られた。これらの解析に加え、一昨年度から花の匂いの比較解析も行ってきた。本年度はイヌビワとイヌビワコバチ、オオバイヌビワとオオバイヌビワコバチの種内の集団間の遺伝的分化ならびに植物の花の匂いの違いを調べた。その結果からも分かるように、種間において必ずしも同様なプロセスで遺伝的分化が起きるとは限らない。イヌビワとイヌビワコバチは集団間で互いに類似した遺伝的分化が起きており、有意な相関も見られる。一方、オオバイヌビワとオオバイヌビワコバチでは、植物のほうは遺伝的にも花の匂いも明白に集団間の分化が起きているが、コバチの方は、それに対応した分化が見られない。イチジクとイチジクコバチの厳密な共生関係を考えると興味深い知見が得られたといえる。今後の調査と解析を進めることによって、イチジクとイチジクコバチの種分化のメカニズムの解明に繋げて行きたい。