年度別活動報告
年度別活動報告書:2008年度
分子系統から生物進化を探る 3-1.イチジク属植物とイチジクコバチの共生関係と共進化
蘇 智慧(主任研究員) 楠見淳子(奨励研究員)
山崎雅美、松浦純子、尾川武史、佐々木綾子(研究補助員) 石渡啓介、魚住太郎、上田千晶(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの相利共生関係の構築、維持、崩壊のメカニズム、種分化の様式などについて分子系統解析や集団遺伝学的解析などにより研究を行っている。
イチジク属Ficusは、クワ科 (Moraceae) に属し、4亜属 (イチジク亜属Ficus, アコウ亜属Urostigma, Pharmacosycea, Sycomorus)に分けられ、東南アジア、アフリカ、中南米など熱帯を中心に約750種が世界中に分布している。日本はイチジク属の分布域の北限にあたり、南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。そのうち、小笠原諸島に固有種3種と移入種1種がいる。一方、イチジク属植物に共生、或いは寄生している一群のイチジクコバチの仲間は、分類学的に膜翅目のコバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとされている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つが送粉コバチ亜科Agaoninae (fig-pollinating wasps)で、他の亜科はすべて非送粉コバチによって構成されている。送粉コバチ類は同定されているものだけで20属300種以上あり、植物と同様に種多様性が高い。
イチジク属植物と送粉コバチとの間、子孫を残す共通の目的のもとで、「1種対1種」関係が結ばれていると言われている。この「1種対1種」関係を維持しながら種分化が起きるとしたら、同調した種分化や系統分化が起こることが予想される。これまで、分子系統学的解析を用いてこの仮説を検証する研究が行われてきた。その結果、イチジク属の節(section)レベルの系統関係とコバチ類の属のそれとがおおまかに一致し、仮説が支持されるものの1)、種間、種内レベルでは系統関係の矛盾のほかに、送粉コバチの隠蔽種や1種のイチジク属植物に複数種の送粉コバチが共生するなど、不明瞭な点が残っている2-4)。そこで、我々は、本年度主として種内レベルの遺伝的多様性、集団間の遺伝的分化の程度を集団遺伝学的手法により解析し、それらがイチジク属植物と送粉コバチ類で同調しているかを明らかにすることで、仮説の検証を試みる。さらに、生物間相互作用は生物の適応進化を促すと考えられ、環境要因と同様、生物多様性を生み出す原動力のひとつとなっている。生物間の相互作用が、種分化の発端となる生殖的隔離の原因となり、両種の種分化が促進されたかは興味深い問題である。イチジク属植物と送粉コバチ類の種分化プロセスの遺伝的背景を明らかにすることで、共進化と種分化、そして種多様性との関わりについて考えてみたい。本研究では、日本(本州関西以西)を北限とし、熱帯、温帯アジアに広く分布しているイヌビワ(Ficus erecta)、そしてその近縁種であり小笠原諸島固有種であるトキワイヌビワ(F. boninsimae)、オオトキワイヌビワ(F. nishimurae)、それに共生するコバチ類(Blastophaga japonica, Blastophaga sp.)を材料とする。
結果と考察
1)マイクロサテライトマーカーの開発
種内多型の解析には、多型性の高いマイクロサテライトマーカーを用いた解析が適しているので、本年度はイヌビワを中心に近縁種の既存のマーカーの配列を基にマイクロサテライトマーカーの開発を行ない、10個のマーカーを作成した。このうち5個のマーカーはイヌビワとトキワイヌビワの両種で共通して解析可能であり、2個はイヌビワのみ、3個はトキワイヌビワとオオトキワイヌビワの解析に用いることができる。今後はイヌビワコバチのマーカーの開発を進める。
2)イヌビワとイヌビワコバチの遺伝的多様性、集団間の遺伝的分化の比較解析
イヌビワで利用できる7個のマーカーを用い、日本(大阪、琉球諸島、八重山諸島)、台湾のイヌビワサンプルの解析を行った。その結果、台湾集団と3つの日本集団のイヌビワには有意な遺伝的分化があることが示された。従って、イヌビワでは地理的に離れた集団間での交配(遺伝子の流動)はある程度制限されていることが予想される。
3)イヌビワ、トキワイヌビワ、およびそれに共生するコバチの種分化
典型的な海洋島である小笠原諸島には、大陸から分布を拡大した生物が島内で独自に進化を遂げた固有種が多くみられ、イチジク属も3種の固有種(トキワイヌビワ、オオトキワイヌビワ、オオヤマイチジク)が生息している。葉緑体DNAの塩基配列に基づいた分子系統学的解析結果では、小笠原の固有種と、祖先種とされているイヌビワとの間に起こった塩基置換は非常に少なく、系統解析からはこれらが姉妹群となることが支持された。従って、小笠原固有種は、イヌビワもしくはその近縁種を単一の祖先とし、諸島内で種分化したことが示唆される。さらに詳細に小笠原諸島でのイチジク植物とコバチ類の種分化プロセス、分岐年代の推定を行うため、植物の3つの核遺伝子(Tpi, G3pdh, Aco1)、遺伝子間領域(ITS)、そしてコバチのミトコンドリア遺伝子(CytB, CO1, CO2)と28Sの塩基配列の決定、分子系統解析を進めている。材料としては、イヌビワ、トキワイヌビワとその近縁種を用いる。これまでに得られたITS、Aco1遺伝子、送粉コバチの28Sの塩基配列情報に基づき、分子系統解析を行ったところ、当初の予想に反し、Aco1遺伝子の分子系統樹ではトキワイヌビワとイヌビワが姉妹群とならなかった(図2)。また、トキワイヌビワの核のITS領域の塩基配列の中には、イヌビワとは大きく異なる配列をもっているものがみられた(図3)。これらの結果から、トキワイヌビワの祖先系統はイヌビワと別種の種間交雑によってできた雑種ではないかと考えている。従って、小笠原諸島のイチジク属の固有種形成には、地理的要因だけでなく、雑種形成やそれに伴う生物間相互作用の適応進化等の生態的要因が影響している可能性が考えられる。一方で、コバチの28Sの分子系統樹ではイヌビワコバチとトキワイヌビワに共生するコバチが姉妹群を形成し、イヌビワコバチが単一の祖先であることが示唆された(図4)。
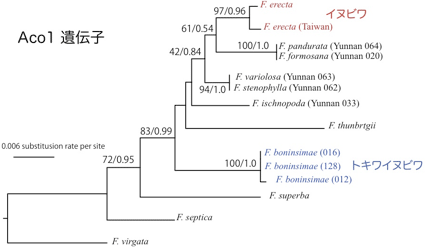 |
図2 Aco1遺伝子によるイチジク属植物の系統樹。最尤法(ML)とベイズ法(BI)により作成した。(枝上の数値はML/BI法の信頼度を表す)
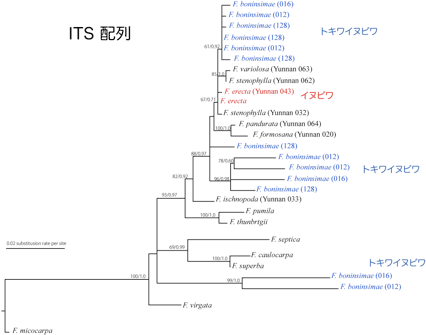 |
図3 ITS配列によるイチジク属植物の系統樹。ML法とBI法により作成した。括弧内の信号は各々のサンプルの番号を示す。トキワイヌビワでは一つの個体から塩基配列が大きく異なる配列が得られた。(枝上の数値はML/BI法の信頼度を表す)
 |
図4 28SrRNA遺伝子による送粉コバチ類の系統樹。ML法とBI法により作成した。(枝上の数値はML/BI法の信頼度を表す)
おわりに
イチジク属とイチジクコバチとの共進化と共種分化のメカニズムを解明するためには、集団内の遺伝的多様性と分化および近縁種間の関係の解明がキーポイントの1つである。本研究は、これまで分からなかったイヌビワの地域的分化を明らかにした。しかし、図1に見られるように、サンプル数がまだ少なく、特に本州のサンプルを増やす必要がある。また、イヌビワの送粉コバチには植物と同調した地域的分化が起きているかどうかも解明する必要がある。一方、小笠原諸島の固有種の種形成に関しては交雑が関与していることが示唆されたが、さらなる分子遺伝的証拠の提示と、できればどの種が交雑に関わっていたかを明らかにしたいと考えている。