年度別活動報告
年度別活動報告書:2007年度
3.分子系統から生物進化を探る 3-3.植物の多様化と遺伝子の多様化
蘇 智慧(研究員、代表者) 佐々木剛(奨励研究員)
石渡啓介(大阪大学大学院生) 魚住太郎(大阪大学大学院生)
神田嗣子(研究補助員) 山﨑雅美(研究補助員)
松浦純子(研究補助員)
はじめに
これまでに動物の多数の遺伝子族において、機能・構造の異なる遺伝子の多様化は形態の多様化に先行して完了していたことが明らかにされてきた。このことから、形態の多様化は新たな遺伝子を作ることによってではなく、既存の遺伝子を利用することによって達成されたことが考えられる。このことを一般化する目的で、植物の多様化と遺伝子の多様化の関連を明らかにする研究を宮田顧問と共同で進めた。
結果と考察
細胞間情報伝達や発生制御に関与する受容体型キナーゼ (receptor-like kinase; RLK) 遺伝子族の多様化パターンを分子系統進化学的に解析した。コケ植物のゼニゴケ、接合藻類のミカヅキモ、シャジクモ目のジュズフラスコモから、PCR法を用いてRLK遺伝子を探索した結果、ゼニゴケから29個、ミカヅキモから14個、ジュズフラスコモから13個のRLK遺伝子を単離できた。単細胞藻類のミカヅキモに多細胞特有のRLK遺伝子が複数見つかったことは非常に興味深い結果である。さらに、これらの遺伝子について細胞外受容体ドメインを含むcDNAの全配列決定を行った。その結果、ゼニゴケの24遺伝子、ミカヅキモの6遺伝子、ジュズフラスコモの8遺伝子が受容体型であった(図3, 4)。ゼニゴケの配列データを加えてRLK遺伝子族の分子系統樹を推定した結果、細胞外ドメインなどの構造の異なるサブファミリーの多様化(重複およびドメインシャフリング)はゼニゴケと維管束植物の分岐以前にほぼ完了していたことが明らかとなった。同じサブファミリーに属する遺伝子の重複は、コケ植物と維管束植物の分岐前にも後にも起きていた。さらに、ミカヅキモおよびジュズフラスコモのRLK遺伝子も加えた解析により、サブファミリーの多様化をもたらした遺伝子重複の多くは陸上植物と接合藻類との分岐やシャジクモ目との分岐より以前にまでさかのぼることも明らかになった(図5)。以上の結果から、形態の多様化に先立ってサブファミリーの多様化が完了していたという遺伝子の多様化パターンは、動物だけでなく植物にも当てはまる可能性が示唆された11)。
おわりに
今後、接合藻類のゲノム配列解析を行うことによって、植物の多数の遺伝子族において同様の多様化パターンが見られるかどうかの解析が可能になり、形態の多様化が既存の遺伝子の利用により達成されたという仮説がより一般化されることが期待できる。
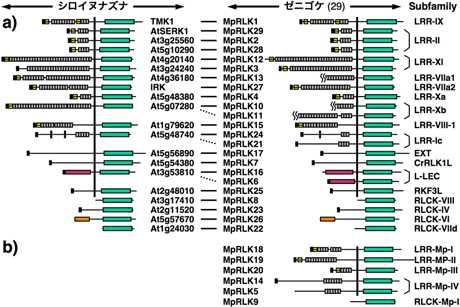 |
図3. A) シロイヌナズナに同じサブファミリーのメンバーが存在するゼニゴケのRLK遺伝子。B) ゼニゴケ特有のRLK遺伝子。
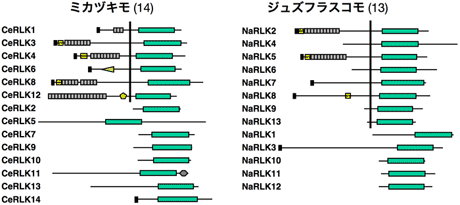 |
図4. 接合藻類のミカヅキモおよびシャジクモ目のジュズフラスコモから単離したRLK遺伝子。
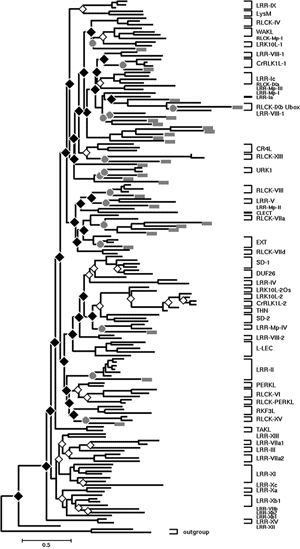 |
| 図5. RLK遺伝子族のキナーゼドメインによる分子系統樹。シャジクモ目ジュズフラスコモおよび接合藻類ミカヅキモの遺伝子を灰色長方形で、それぞれの藻類と陸上植物との分岐を灰色の丸で示した。また、この分岐以前に起きた遺伝子重複を黒の菱形で、時期不明の遺伝子重複を白の菱形で示した。 |
引用文献
1) Weiblen G.D. (2000) Phylogenetic relationships of functionally dioecious Ficus (Moraceae) based on ribosomal DNA sequences and morphology. American Journal of Botany 87: 1342-1357.
2) Machado C.A., Robbins N., Gilbert M.T.P. and Herre E.A. (2005) Critical review of host specificity and its coevolutionary implications in the fig/fig-wasp mutualism. Proc. Natl. Acad. Sci. USA 102 (Supplement 1): 6558-6565.
3) 蘇智慧, 飯野均, 中村桂子 (2004) メキシコ産イチジクコバチの分子系統と進化. 蝶類DNA研究会ニュースレター No. 12: 39-43.
4) Su Z.-H., Iino H., Nakamura K. Serrato A. and Oyama K. (2008) Breakdown of the one-to-one rule in Mexican fig-wasp associations inferred by molecular phylogenetic analysis. Symbiosis 45:73-81.
5) 東浩司, 蘇智慧, 中村桂子 (2006) 日本産イチジク属各種とその送粉コバチの分子系統関係. 昆虫DNA研究会ニュースレター, No. 4: 6-15.
6) Nardi F., Spinsanti G., Boore J.L. et al. (2003) Hexapod origins: monophyletic or paraphyletic? Science 299: 1887-1889.
7) Rejier J.C., Shultz J.W. and Kambic R.E. (2005) Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic. Proc. R. Soc. B 272: 395-401.
8) Braband A., Richter S., Hiesel R. and Scholtz G. (2002) Phylogenetic relationships within the Phyllopoda (Crustacea, Branchiopoda) based on mitochondrial and nuclear markers. Mol. Phylogenet. Evol. 25: 229-244.
9) Stenderup J.T., Olesen J. and Glenner H. (2006) Molecular phylogeny of the Branchiopoda (Crustacea)-multiple approaches suggest a 'diplostracan' ancestry of the Notostraca. Mol. Phylogenet. Evol. 41: 182-194.
10) Olesen, J. (1998) A phylogenetic analysis of the Conchostraca and Cladocera (Crustacea, Branchiopoda, Diplostraca). Zool. J. Linn. Soc. 122: 491-536.
11) Sasaki G., Katoh K., Hirose N., Suga H., Kuma K., Miyata T. and Su Z.-H. (2007) Multiple receptor-like kinase cDNAs from liverwort Marchantia polymorpha and two charophycean green algae, Closterium ehrenbergii and Nitella axillaris: Extensive gene duplications and gene shufflings in the early evolution of streptophytes. Gene 401: 135-144.