年度別活動報告
年度別活動報告書:2006年度
3.分子系統から生物進化を探る 3-1.昆虫と植物との共生関係、共進化および種分化に関する研究
蘇 智慧(研究員、代表者)
佐々木剛(奨励研究員)
神田嗣子(研究補助員)
石渡啓介(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの相利共生関係の構築、維持、崩壊のメカニズム、種分化の様式などについてDNA系統解析や野外観察調査などにより研究を行っている。
イチジク属Ficusは、クワ科 (Moraceae) に属し、4亜属 (イチジク亜属Ficus, アコウ亜属Urostigma, Pharmacosycea, Sycomorus)に分けられ、東南アジア、アフリカ、中南米など熱帯を中心に約750種が世界中に分布している。日本はイチジク属の分布域の北限にあたり、南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。そのうち、小笠原諸島に固有種3種と移入種1種がいる。一方、イチジク属植物に共生、或いは寄生している一群のイチジクコバチの仲間は、分類学的に膜翅目のコバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとされている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つが送粉コバチ亜科Agaoninae (fig-pollinating wasps)で、他の亜科はすべて非送粉コバチによって構成されている。
イチジク属植物と送粉コバチとの「1種対1種」の関係は古くから共進化のモデル系として多くの研究がなされてきた。近年において分子系統解析の研究も盛んに行われるようになった。これまでの報告によると、イチジク属植物と送粉コバチとの系統関係は、分類上のイチジク属のsectionとコバチのgenusと大まかに対応していることが示唆されている。この結果は、基本的にイチジク属植物と送粉コバチとの協調的種分化と共進化を支持している1)。しかし、いくつかの問題点があると私たちは考えている。イチジク属各種がそれぞれ広い分布域を持っており、日本産の大部分の種も東南アジア、オーストラリアまで広く分布している。これほど広い分布域をもち、海や山など大きな地域隔離を有する生物種の系統関係を調べるには地域性を考慮しなければならない。1地域或いは1サンプルだけで種の系統的位置を決めるのはかなり危険であり、異なった結論に導く可能性がある。実際、最近送粉コバチの隠蔽種や、1種のイチジクに複数種の送粉コバチの存在などのことも報告され始めている2)。従って、「1種対1種」の関係を検証するためにも、分布域を網羅する複数地点からの送粉コバチを分析する必要がある。しかし、これまでの報告を見ると、このような緻密な解析はほとんどなされていない。
我々は以上の問題点を考慮した上、これまでメキシコ産、日本産と中国海南島産のイチジクとイチジクコバチの分子系統解析を行ってきた。メキシコ産の材料を分析したところ、Urostigma亜属の送粉コバチはおよそ2100万年前に一斉放散分化したことが示唆され、また、同じ種のイチジクから採集した送粉コバチが系統樹上1つの枝にまとまらず、異なる系統に分かれるという興味深い結果が得られた(原著論文投稿中)3, 4)。この結果はイチジク属植物と送粉コバチの「1種対1種」関係の崩壊を示唆した。とことが、日本産の材料を調べたところ、その「1種対1種」の関係はほぼ厳密に維持されている(アコウとその送粉コバチを除く)ことが分かった。また、昨年度は葉緑体DNAと核ITSを使い分けにして、日本産イチジク属の上位分類群間の系統関係も明らかにした(原著論文作成中)5)。今年度は中国産イチジク属植物と日本産のものとの関係の解明を中心に研究を進めてきた。
結果と考察
1) 中国産イチジク属植物とコバチの系統解析
まず植物の系統解析はこれまでと同様、葉緑体DNAの6つのイントロン部分の塩基配列と核ITS領域を用いた。ITSは核ゲノム上に、複数コピーがあるので、クローニングして、1サンプルに対して5-10クローンを取って塩基配列の決定を行った。乾燥葉から全DNAを抽出し、PCR反応によって目的のDNA断片を増幅し、塩基配列を決定した。一方、コバチの方は、核28SrRNA遺伝子とミトコンドリアCOI遺伝子を使用した。コバチは体長が1-2mm くらいしかない小さな昆虫のため、個体全体を使って DNA抽出に用いた。系統解析は、MAFFTというコンピューターソフトウェアによって塩基配列をアライメントし、PAUPとMOLPHYでNJとMLの系統樹を作成した。
図1は中国産と日本産を含むイチジク属植物(左図)とそれらの送粉コバチ(右図)の系統樹である。まず植物の系統樹を見てみよう。幾つかのグループが形成された。日本産のムクイヌビワ、ホソバムクイヌビワとハマイヌビワ3種を含むSycidiumセクション(イチジク属の分類単位)と、ガジュマルを含むConosyceaセクションはそれぞれ単系統を形成することが分かった。これら2セクションの各種から得られた送粉コバチは、系統樹上の信頼度が高くないが、それぞれ2つのグループにまとまっている(図1の右図を参照)。また、日本産と中国産のギランイヌビワと他の中国産2種 (F. auriculata, F. oligodon hainanensis) を含むNeomorpheセクションと、日本産アカメイヌビワとオオバイヌビワおよび中国産2種 (F. hispida, F. harlandii) を含むSycocarpusセクションが共通祖先を有していることは系統樹上高い支持率 (96%) で示唆された(左図)。これら2つのセクションの各種から採集された送粉コバチも92%の高い信頼度で共通起源であることが判明した(右図)。これらの結果は、イチジク属植物とイチジクコバチの同調的種分化および共進化説を支持している。ところが、Sycidiumセクション、RhizocladusセクションとFicusセクションの一部は同一グループを形成しているが、それらの送粉コバチは同じグループにまとまらず、独立した系統に分かれている。従って、イチジク属植物とイチジク送粉コバチとの共生関係において、同調的種分化と共進化は実際に存在していることが判明されており、極めて重要な考え方ではあるが、そうでない部分もあることを同時に考慮に入れて両者の関係を理解する必要がある。
2) イチジク属植物とその送粉コバチの平行進化
日本産のイチジク属には、イタビカズラ (F. sarmentosa var. nipponica) とヒメイタビ (F. sarmentosa var. thunbergii) という蔓性の種がいる。前者は日本の本州(福島・新潟以西)、四国、九州、沖縄、それから、朝鮮半島、中国南部、台湾に分布しているが、後者は日本の本州(千葉県以西)、四国、九州、沖縄と韓国の済州島に分布域を持っている。日本産のこれら2変種は極めて近縁で、形態的に辛うじて判別できるが、分子系統樹上区別つかない(図1参照)。
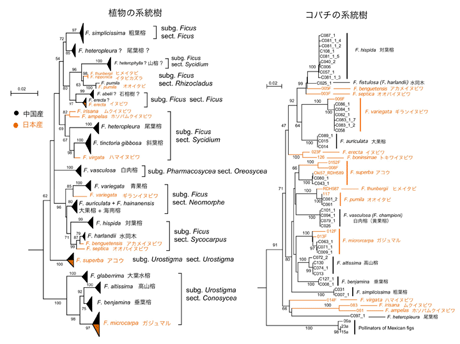 |
図1. 核ITS領域による中国産と日本産イチジク属植物の系統樹(左図)と28SrRNA遺伝子による送粉コバチの系統樹(右図)。枝の分岐点にある数字はBootstrap確率で、系統樹の信頼度を示している。比較しやすいため、コバチの系統樹にも植物の学名を当てている。
日本産イチジク属とその送粉コバチの起源と地域変異の解明の一環として、今年度の夏、中国雲南省の西双版納で開催されたイチジク属植物とイチジクコバチの国際シンポジウムに出席した際、運良くF. sarmentosaとその送粉コバチを採集することができた。早速サンプルからDNA抽出し、系統解析を行ってみたが、思い掛けない結果となった。中国産のF. sarmentosaは日本産のものと全く近縁性を示さず、完全に異なった別系統であることが分かった。さらに興味深いことは、この中国産のものは全く別種のF. pubigeraと組み、日本産のものはまた別種のF. pumilaと組んでいた(図2)。また、これら3種の送粉コバチの系統解析を行ったところ、植物と同じ系統関係が示された(F. pubigeraの送粉コバチは残念ながらまだ採集出来ていない)(図3)。この結果はF. sarmentosaとその送粉コバチが平行進化したことを強く示唆した。
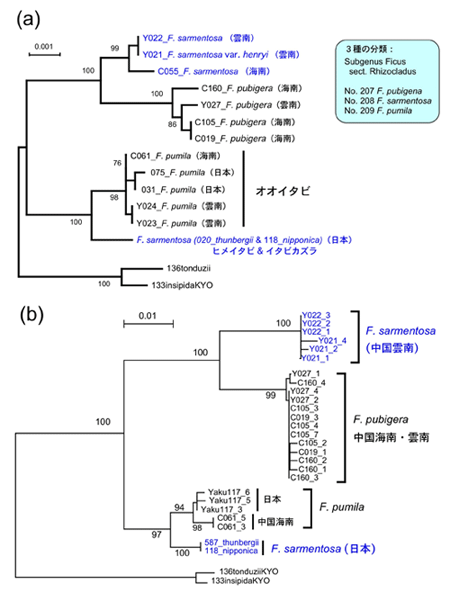
図2. 葉緑体DNA (a) とITS (b) による3種のイチジク属植物の系統樹。
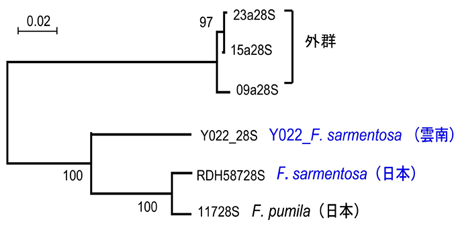 |
| 図3. 28S rDNAによるF. sarmentosaとF. pumilaの送粉コバチの系統樹。学名は植物の名前を示している。 |
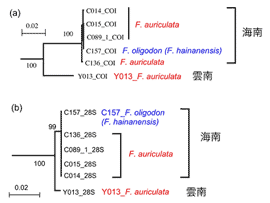
上述と同様な現象は中国産イチジクF. auriculataにも観察された。この種はヒマラヤから中国南部にかけて、インド北東部、タイ、インドシナまで広く分布している。非常に大きな花嚢と葉を持つことが特徴である(図4)。これまでの解析では、中国海南島で採集されたこの種は同じく海南島にいるF. oligodon (F. hainanensis) という種と極めて近縁であることが判明されている。今年度の夏、中国の雲南省からもこれら2種およびF. auriculataの送粉コバチを採集することができた。系統解析を行ったところ、これもまた驚いた結果となった。種ごとにまとまらず、地域系統に分かれた。つまり、海南島産のF. auriculataとF. oligodon、雲南産のF. auriculataとF. oligodonがそれぞれ近縁で、両地域が分かれることが示された(図5)。コバチの系統解析も同様な結果が得られた(図6)。これらのことも両種がそれぞれの地域で平行進化したことを示唆している。
 |
図4. Ficus auriculata (中国海南島産)。
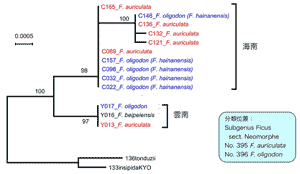
図5.葉緑体DNAによる植物の系統樹。平行進化を示す。
おわりに
イチジク属とイチジクコバチの関係は寄生コバチを含めた三者の関係が絡み合った複雑な系である。イチジク属と送粉コバチの共生関係の進化を探る上ではこれら三者の生活史がどのように関連しているかを知る必要があるが、まだほとんど分かっていないのが現状である。特に、非送粉コバチの大部分については、種の同定さえできていない。今年度はイチジク属植物とイチジクの平行進化など興味深いことを発見したので、この結果の確実性を高めるために、まだ採集されていない必要な送粉コバチを入手して解析したいと思っている。また、非送粉コバチの系統関係、中国産イチジク属植物とイチジクコバチの詳しい系統関係および日本産のものとの繋がりなどを中心に解析を行い、イチジクコバチによる宿主転換の可能性や転換機構および種分化における生態的なメカニズムに関する理解を深めたい。
 |
|
図6. ミトコンドリアCOI遺伝子 (a) と28S rDNA (b) による送粉コバチの系統樹。学名は植物の名前である。
|