年度別活動報告
年度別活動報告書:2000年度
DNA による昆虫類の多様化に関する研究 1. DNA による昆虫類の多様化に関する研究 (1) オサムシの分子系統と進化 ---- 論文作成を中心にして (2) ゴマダラカミキリ属の進化
蘇 智慧(研究員)
大澤省三(顧問)
飯野 均(阪大連携大学院生)
岡田節人(館長)
中村桂子(副館長)
はじめに
昆虫は現在最も多様化している動物群で, 生物の多様化(進化)の要因解明するための格好の材料である。また生物進化研究の前提条件は生物の系統関係を明らかにすることである。DNA による系統解析は客観性, 正確性などにおいて従来の形態による系統分類より遙かに優れていることは, 既にオサムシを含む様々な生物の分子系統解析研究によって証明されている。1994 年に開始したオサムシ研究プロジェクトは昨年度一応の終了を迎えた。約 6 年間の研究で, オサムシの一斉放散, 平行放散進化, 地史との関連, 不連続的形態変化, 及び進化における「動」と「静」など数々のことが解明され, 生物の進化の原理を探る上でも極めて重要な知見が得られたと考える。これら膨大な研究成果を論文の形で公表することは重要な研究の一環であり, これによってこれまでの研究成果を生かした新たなステップを見いだすことができるであろう。オサムシ進化の全体像は, これまで正式に国際誌に公表していなかったが, 遠からず Journal of Molecular Evolution に印刷される (Su et al., 200l, in press) ので, その反響が期待される。今年度はオサムシの研究成果を論文に纏めることを中心に, 必要に応じてミトコンドリア遺伝子や核遺伝子を用いた追加分析を行ってきた。またオサムシの研究で得られた興味深い現象の一般化のため, 自然環境センターの斉藤秀生氏及び愛媛大学の小川次郎氏と共同でカミキリムシなど他の昆虫の解析も行った。
結果と考察
トレハラーゼ遺伝子によるオサムシ亜科 (Carabinae) 4族間の系統関係
オサムシ亜科 (Carabinae) は,セダカオサムシ族 (Cychrini) とオサムシ族 (Carabini) に分けられ,オサムシ族はさらにカタビロオサムシ亜族 (Calosomina) とオサムシ亜族 (Carabina) に分割されている。mtND5 遺伝子による系統解析の結果は上記の高次分類を支持している。また Carabinae に属するチリオサムシ (Ceroglossus) とオーストラリアオサムシ (Pamborus) に関しては,核 28S rDNA による解析からそれぞれ独立の系統であり,別族 (Tribe) を形成していると考えられた (1999 年度報告書)が, これら4族間の系統関係にはなお不明確な点が残されていた。そこで我々は昆虫の血糖であるトレハロースを分解する酵素トレハラーゼ遺伝子を解析し, オサムシ亜科4族間の系統関係を調べた。その結果, セダカオサムシ族は他の3族とかなり異なった配列をもち, また他の3族が共有している2個のイントロンも欠失していることが判明した。またチリオサムシ族とオーストラリアオサムシ族を形成する共通クラスターは比較的高い Bootstrap 値で支持された。以上の結果から, セダカオサムシ族は他の3族の共通祖先とかなり古い時期に分かれ, 次いでオサムシ族がチリオサムシ族とオーストラリアオサムシ族の共通祖先と分岐し, チリオサムシとオーストラリアオサムシは祖先を共有していたと結論した(図 1)。
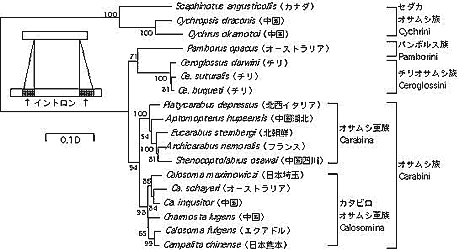 |
【図1】Trehalase 遺伝子によるオサムシ亜科4族の系統関係。
チリオサムシのカラーパターンの同所的収斂進化
上に述べたようにチリオサムシ (Ceroglossus) が独立した族 (Tribe) を形成している。この族は, 多種多様な形態分化を遂げて北半球に広く分布しているオサムシ族に比べ, その生息地は南米のチリとアルゼンチンの一部に限られ基本形態も互いによく似ている。しかし, さまざまな色彩変異が多く見られ, それがチリオサムシの分類を混乱させてきた。最近 Jiroux (1996) が主に触角の微毛の生え方や♂交尾器の構造などを基準にして, チリオサムシを 4 種群, 8 種に分けた。この分類体系によれば, 同所的に生息する別種が色彩的に区別が付かないほど似ている, いわゆる同所的収斂現象が見られることを意味する。今回われわれはチリ各地から採集した 66 個体の mtND5 遺伝子を分析しチリオサムシの系統関係を調べた。その結果, チリオサムシは4 つの系統に明確に分かれ, Jiroux (1996) の分類と完全に一致した。これらの系統は 2500-3000 万年前に分岐したことも推定された。一方, 各系統内には多様な地域に依存した色彩変化がみられる。 この事実はチリオサムシのカラーパターンが種特異的ではなく, 地域特異的であることを意味し(図 2), 色彩変化は生態的ー環境的要素との共進化の結果であることを示唆している (Okamoto et al., J. Mol. Evol., 200l, in press)。
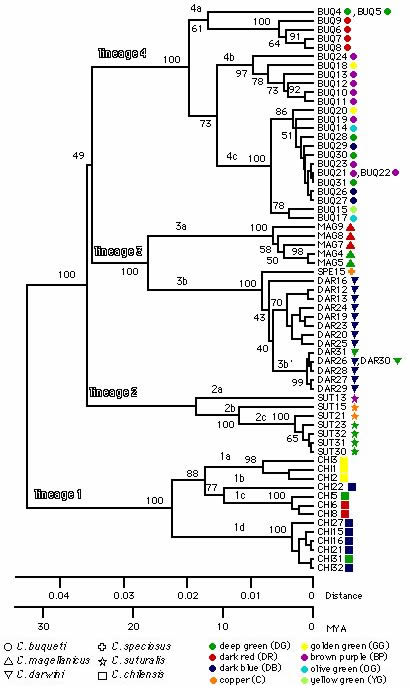 |
【図2】mtND5 遺伝子によるチリオサムシの系統関係
マルオサムシ群の系統関係
マルオサムシ群 (Archicarabomorphi) はヨーロッパから小アジア半島, さらに天山山脈にかけて分布し, 4 亜属からなっており, 15 種ほど知られている。マルオサムシ群を構成する4亜属のうち,3亜属から代表的な種を選び,ミトコンドリア ND5 遺伝子の塩基配列を決定して,群内の系統関係について検討した。分析した3亜属のうち,天山山脈のキンマルオサムシ (Acrocarabus) はもっとも分岐が早く,全体のオサムシと共に系統樹を作成したところ, この亜属のものは骨片オサムシ群に近縁であることが判明した。また♂交尾器内袋に指状片を有するなど,形態学的にもかなり特異なので,マルオサムシ群から除外して骨片オサムシ群に分類すべきものと思われる。北西イランに孤立分布するキバヒョウタンオサムシ (Gnathocarabus) もかなり独立性の高いクラスターを形成したが,分子・形態双方の所見から,マルオサムシ群の一員と考えてよいだろう。狭義のマルオサムシ (Archicarabus) は大きく2サブクラスターに分かれ,一方はトルコ産の2種,他方はヨーロッパ産の2種によって構成された。前者に対しては (Deuterocarabus) 亜属を適用すべきかもしれない。 (Archicarabus) の基準種である nemoralis の塩基配列には産地による相違がほとんどみられず,北米や中央アジアに分布する集団は,従来の指摘どおり,移入種である可能性の高いことが示唆された (Imura et al., 2000)。
ヒラタオサムシ群の分子系統
ヒラタオサムシ群 (Arciferi) は,ヒラタオサムシ亜属 (Platycarabus),キバナガヒラタオサムシ亜属 (Chaetocarabus), ヒメツヤヒラタオサムシ亜属 (Heterocarabus), ミズベオサムシ亜属 (Hygrocarabus) の4亜属からなり,♂交尾器内袋基部に弓状片arculus とよばれる器官を共有することにより,他の群から形態学的に識別される。これら各亜属から1~2種ずつを選び,ミトコンドリア ND5 遺伝子の塩基配列を決定して,群内の系統関係を検討したところ,ミズベオサムシ亜属のもののみは他の3亜属から類縁が遠く,オサムシ亜族 (Carabina) の一斉放散の時期に分岐した独立系統を形成している。この亜属については分類学的に独立した地位を与えるべきものであろうと考えられた。他の 3 亜属も互いに分岐がかなり古いが, オサムシ亜族においては単一の系統を形成している (Imura et al., 2000)。
セアカオサムシ群の系統関係
セアカオサムシ群 (Crenolimbi) はセアカオサムシ亜属 (Hemicarabus) とセスジアカガネオサムシ亜属 (Homoeocarabus) の 2 亜属のみにより構成される最も小さい群だが,その分布は広く,全北区に及ぶ。セアカオサムシ亜属はユーラシアと北米の両大陸に分布し, 4 種を含む。一方セスジアカガネオサムシ亜属はセスジアカガネオサ 1 種のみからなっており, シベリア東部, モンゴル北部, 中国北東部, 韓国済州島, 北海道, カムチャツカ半島, 北米大陸北部に分布している。今回は,本群に属する5種すべてを用いて, ミトコンドリア ND5 遺伝子の塩基配列を決定し,群内における系統関係を検討した。その結果,Homoeocarabus は Hemicarabus から明確に区別されること, Hemicarabus 内においては, ユーラシアに分布する3種は互いに近縁で, 北米の serratus のみ類縁が遠いこと,などが判明した (Su et al., 2000)。
カザリオサムシ群の系統関係
カザリオサムシ群 (Lipastromorphi) は主として中型ないし小型の種から構成される群で, ユーラシア大陸のほぼ全域にわたって繁栄しているが, 中国南部や日本列島は分布の空白域となっている。6 亜属を含む。今回は, ほぼ全分布域から 6 亜属, 40 種を含む 85 個体の mtND5 遺伝子の塩基配列を解析し, カザリオサムシ群内の系統関係を調べた。その結果, カザリオサムシ群はまず 2 つの系統に分かれ, またそれぞれ 2 と 9 亜系統に分岐した。これらの亜系統は地域性を大きく反映していることが判明した(図 3)。Cyclocarabus, Lipaster, Mimocarabus 3 亜属はそれぞれの亜系統を形成しているが, Ophiocarabus 亜属と Cryptocarabus 亜属は同一の亜系統を形成し, また後者は前者の幾つかのグループの間に入っていた。これは Cryptocarabus-type が Ophiocarabus-type から急速な形態変化を起こして分かれてきたことを示唆している。またこれまで Morphocarabus 亜属として分類されてきたものは 7 つの独立した亜系統に分かれ単一のクラスターを形成しなかった。つまりこの亜属は多系統であることが分かった。しかし, これらの亜系統間の基本形態はほとんど変化していない。およそ 3000 万年前から分岐したことから, この亜属はかなり長い進化的“静”の時期を保っていることが考えられる (Imura et al., 2001, Ann. Ent. Soc. Amer., in press)。
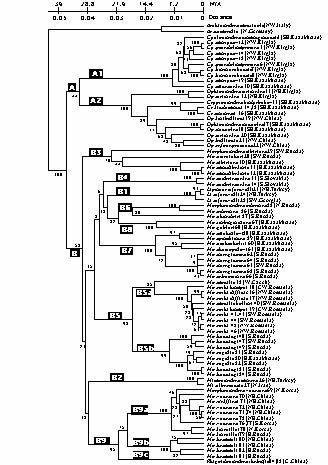 |
【図3】mtND5 遺伝子によるカザリオサムシ群の系統関係
シンポジウム<DNAで辿る昆虫の系統と進化>について
去る平成 12 年 2 月 11 日~12 日, BRH 主催のシンポジウム「DNA で辿る昆虫の系統と進化」が大阪府高槻市のたかつき京都ホテルで開催された。1999 年度の報告書に間に合わなかったため今年度の報告書に報告したい。このシンポジウムについて北海道大学農学部昆虫体系学講座の溝田浩二氏が「月刊むし」に「シンポジウム・DNA で辿る昆虫の系統と進化に参加して」を題とした紹介感想文を書かれている(月刊むし, No. 352: 42-44, 2000)。ここではその感想文を引用しながら報告したい。
<オサムシ研究グループは, オサムシが歩んできた進化の道筋を再現しようという壮大なテーマのもと, 1994年以来徹底的に世界中のオサムシの DNA 解析を行ってきた。分子系統学の急速な発展, コンピュータの高速化といった時代の流れにも後押しされて, 脅威的なスピードでオサムシの進化過程を解き明かし, “平行放散進化説”などといった眼からウロコの進化理論の枠組みを提出してきた。ひとつの生物グループが, 分子系統学の立場からこれほどまとまって解析されたのは, おそらく世界でも初めてのことだろう。今回のシンポジウムは約6年にわたる BRH のオサムシ研究プロジェクトの終焉と, その総括の意味を込めて催されたものであった。
シンポジウムの初日, 大ホールに用意された机座席は満席になるほどの盛況ぶりで, 会場の両端, 後方の椅子座席もおおよそ埋め尽くされた。予想をはるかに上回る 223 名もの聴衆が関西地方を中心に全国各地から集まり, 今回のシンポジウムに寄せる関心の高さを改めて感じた次第である。今回のシンポジウムの演者は, BRH のオサムシ研究チームはもとより, 高校教師, 大学院生, 大学教官, 医者, など実に多彩な職業・立場の方々である。多様な生き物の世界を調べるのであるから, 調べる側が多様であるのも十分うなずける話であるが, いずれも BRH のオサムシ研究に触発されて, 自らも DNA 解析を始めるようになったという根っからの“虫好き”ばかり。材料も, オサムシを筆頭に, クワガタムシ, カミキリムシ, チョウ, トンボ, ハエ, フナムシ・・・と多種多様であった。 BRH が毎年夏休みに開いている「DNA 実習サマースクール」への参加が縁となっている方もあり, BRH 側の地道な努力が実を結んだ結果ともいえよう。「生命誌」という視点からプロとアマチュアを, 形態と分子を, そして科学と社会とを橋渡しする形で研究を進めてきた BRH の姿勢を見事に象徴しているのではなかったろうか。
午後1時にシンポジウムはスタート。まず岡田節人 BRH 館長からユーモア溢れる挨拶があり, これまで協力していただいた多くの方々への感謝の言葉が述べられた。特に心に残ったのは, 「今回のシンポジウムをもって BRH のオサムシ研究プロジェクトは一応の完結をみましたが, 学問には終わりはなく, “わかった”とか“終わった”とかいうのは幻想でしかありません」いうお話。その言葉どおり, 今回のシンポジウムは新しい進化理論を構築していく過程での第一歩の終わりであり, また新たなステップの始まりであるということを予告しているように思われた。
続いて, 大澤省三氏による基調講演「分子時計が語る昆虫の系統進化」があり, 分子時計のしくみ等を簡潔に解説していただいた。その後, 井村有希氏「分子系統解析に基づくオサムシの高次分類」, 金衝坤氏「ヨロイオサムシ群の分子系統」, 岡本宗裕氏「クビナガオサムシとカブリモドキ周辺の分子系統」, 柏井伸夫氏「チリオサムシの分子系統と収斂進化」, 蘇智慧氏「オサムシの非連続的形態進化」の5題の講演があった。 BRH グループの膨大な分子系統解析の結果は, その内容・データ量ともに圧巻で, オサムシの進化と起源, 生物地理, 形態進化といった多岐にわたる内容について話題を提供された。短い討論の後, 午後6時より懇親会が同ホテルで立食形式で行われた。はじめに行われた挨拶では, 中村桂子 BRH 副館長が「米スペースシャトル・エンデバー号で宇宙飛行中の毛利衛さんに, Carabus (Shenocoptolabrus) osawai (大澤省三氏の名にちなんだ中国四川省産の美麗種オサムシ)を樹脂に封入した形で一緒に宇宙に連れていってもらったんですよ」というエピソードを紹介。飛べないオサムシが宇宙に飛び立ったという粋なはからいに, 会場は拍手喝采だった。乾杯の後, 日本各地からやってきた参加者たちの間でコップを片手に交流の輪が広がった。
翌日は午前9時から, 富永修氏の「日本列島におけるオサムシ相の成立」で再開し, 伊谷結氏「フナムシの分子系統解析─日本産フナムシの系統・島嶼のフナムシ・移入種」, 斉藤秀生・斉藤明子両氏「分子系統解析にもとづくハナカミキリ類の分類学的再検討」, 荒谷邦雄・近雅博両氏「コガネムシ上科の分子と形態にもとづく系統解析」の4題の講演が午前中に行われた。荒谷・近両氏が提出した日本産クワガタムシの分子系統解析の結果は, 成虫の外部形態からは予想もできなかった衝撃的なものだったが, これは幼虫・蛹の形態形質を用いた系統解析の結果とほぼ一致したという。
午後はひき続き, 桝永一宏氏「海浜性アシナガバエの外部形態解析と分子系統解析」, 林克久氏「不均翅亜目数科のトンボの分子系統」, 八木孝司氏「DNA からみたアゲハチョウの系統と進化」, 近藤喜代太郎・新川勉両氏「鱗翅類の分子系統─トリバネアゲハのきた道」の4題の講演があった。世界でもっとも巨大とされるトリバネアゲハの起源を解明するというのは, チョウ屋なら誰しも1度は思い描いたことのある甘美な夢ではなかろうか?
最後に, 大澤, 尾本, 荒谷の3氏が進行役を務めて総合討論がもたれた。広範な専門分野の方々から疑問点や意見が出されたが, 最後に神戸大学の河野和男氏から「BRHのオサムシ研究は, 進化は不連続に起こるということを物質レベルで証明した世界でもはじめての研究例であり, 今後ネオダーウィニズム的な思考には変更が迫られるでしょう」という総括的なコメントがあり, 充実した2日間のシンポジウムを締めくくった。
今回のシンポジウムを契機として, 虫屋と現代生物学の距離がぐっと近くなったように感じられるが, それだけでも参加者の世界観・自然観が広がったことを意味しているのではないだろうか。形態と分子という異なる証拠に基づくアプローチが楽観的にひとつの結果に収束する保証はどこにもなく, むしろ多くのケースで, それぞれは多かれ少なかれ異なる結果を導いてしまうようである。しかし, 系統的類縁と形態的類似が一致しないからこそ分子系統学は面白いのである。分子データと形態データの双方が有機的に結合することによってはじめて, 生物進化のメカニズムを総合的に理解することができるのであろう。 BRH のオサムシ研究に端を発した昆虫分子系統学は急速にほかの分類群にも波及しており, まさに昆虫の系統学, 分類学の新しい時代の到来といってもよいだろう。21 世紀を出発した今「分子系統学」は, 生物の系統と進化について, どんな問題を提起し, どんな展望を与え, 何を物語ってくれるのか ---- 興味津々である。>
ゴマダラカミキリムシ属の系統と起源
ゴマダラカミキリ属 (Anoplophcra) は日本, 中国, 朝鮮半島など東アジアを中心に分布し, 軽微な形態的な差から幾つかの種, 亜種に分けられているが, 互いに酷似していて, その判別は容易でない。日本からは 3 種類が知られ, ゴマダラ (A. malasiaca) は日本本土と沖縄本島に, オオシマゴマダラ (A. oshimana) は鹿児島県奄美大島と沖縄本島に, タイワンゴマダラ (A. macularia) は琉球諸島に, それぞれ分布している。これらの種分化のパターンと起源を探るため, ミトコンドリア COI 遺伝子を用い, 中国とベトナムの近縁種を含めて系統解析を行った。その結果, ゴマダラカミキリ属は大きく3 つの系統(タイワン, オオシマ, ゴマダラ)に分かれることが判明した。タイワンとオオシマの両系統は共通祖先を持つ可能性も示唆された。これらの系統にはベトナム (A. sp.)と上海のもの (A. chinensis) も含まれるが, ベトナムのものはこの系統の外群になる。この系統関係から, オオシマとタイワンの 2 つの系統は古く大陸から入ってきたものが島ごとに分化した系統と考えられ, 日本のゴマダラ (A. malasiaca) は最近中国大陸から朝鮮半島経由で入ったものと思われる。日本本土のゴマダラと沖縄本島のゴマダラは系統樹で一応2つのグループに分かれるが, かなり近縁で, 沖縄のものは本土から進入したと推定される(図 4)。現在, カミキリの COI 遺伝子の絶対進化速度が不明であるが, Anoplophcara の分化開始はかなり古いと推定される。しかし, その形態分化は微少で, オサムシでみられた“静の進化”が, かなり一般的であることを示唆している。
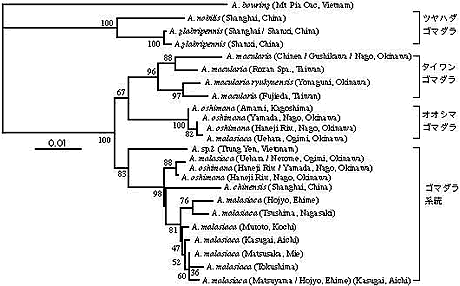 |
【図4】mtCO 遺伝子によるゴマダラカミキリムシの系統関係
おわりに
すでに「はじめに」でも述べたようにオサムシ研究の膨大なデータから生物の多様化に関する様々な知見が得られた。特に進化のパターンについては, ダーウイン以来広く信じられている斬新的進化の考えを支持せず, 進化研究全体に対するインパクトは大きいと考える。これらの成果を順次専門誌などに公表してきたが, プロジェクト終了の昨年度時点で発表に至らなかったデータを整理したところ, まだ 20 数編の論文を書かねばならないことが分かり, その約半数は今年度で完成させた。残りのものを全て論文にし, 最終的には研究書(英・和)の形で纏めるつもりである。岡田館長の話「学問には終わりはなく, “わかった”とか“終わった”とかいうのは幻想でしかありません」のように, オサムシ研究プロジェクトの終了は単なる第一歩の終わりを意味するにすぎない。事実, プロジェクトの終結をむかえた今日でも, まだ詰めねばならないことが山積している。これらを早急に解決するとともに, これまでの成果を基にして, 新たな発展にたち向かいたい。