年度別活動報告
年度別活動報告書:1998年度
DNAによるオサムシの系統解析
蘇 智慧(研究員)
大澤省三(顧問)
金 衝坤(奨励研究員)
岡田節人(館長)
中村桂子(副館長)
はじめに
この地球上で最も繁栄している生物群が昆虫で、動物の総種数の70%以上を占める。従って、このグループは生物の多様化の要因を究明するための絶好な材料である。昆虫の系統分類は従来交尾器を含む形態的特徴によって行われてきたが、主観的要素が入り易く、信頼度は低い。一方、DNA の塩基配列の違いは分岐後の時間経過を示す分子時計とみなすことができ、客観的な系統樹を得ることができる。本研究は形態的データの蓄積が多いオサムシを材料として、分子系統樹を構築し形態的知見と比較しつつ、従来の系統分類を根本的に再検討している。その上に立って生物の多様化(進化)、種分化の要因(特に形態的分化と遺伝子発現の関係、地史との関連)を考察する。これまでオサムシ亜族の一斉放散,オオオサムシ亜属の並行放散進化,マイマイカブリの日本列島地史との関連など数々のことが明らかになった。ここでは今年度の解析によって解明されたことを報告する。
結果と考察
mtND5 遺伝子による年代推定
分子系統樹からの分岐年代の推定は,化石や地史のデータを必要とするのでかなり難しい。だが,年代スケールの設定は系統進化研究の最も重要なポイントの1つで,これがないと系統的データの解釈が大幅に限定されてしまう。
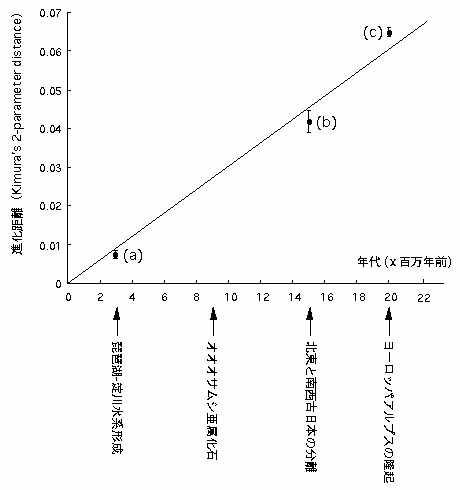
日本特産のアキタクロナガオサムシの亜種ホソアオクロナガオサムシ (Euleptocarabus porrecticollis kansaiensis) は近畿全域に分布するが,淀川を挟んで東側のものと西側のものは mtND5 遺伝子の塩基配列で明らかに区別できる。淀川水系は約 400 万年前にはまだできておらず,約 300 万年前に完成したと推定されているので,この東西 2つの集団は淀川水系の完成による隔離で分岐したと思われる。この 2 集団間の進化距離 (D) は 0.0076±0.0011 と計算されるので,D 0.01 ユニットは 400±50 万年に当たることになる。
もう1つ日本の特産種マイマイカブリ (Damaster bloptoides) は日本列島の形成と関連して系統を形成している。古地磁気の研究から,古代日本は約1500万年前にユーラシア大陸から分離し,東日本と西日本に分かれたとされている。マイマイカブリの祖先もこの時に東系統と西系統に分かれたと推定される。この2系統間の D は 0.042±0.0029 で,上述のアキタクロナガオサムシの年代スケールで換算すると 1680±120 万年になり,地質学のデータとほぼ一致する。
さらに,ヨーロッパに広く分布しているクロツヤオサムシ (Phricocarabus glabratus) はアルプスを挟んで北の集団と,南の集団が mtND5 遺伝子では明確に区別できるので,この2系統はアルプスの隆起による地理的隔離によって形成されたと推定される。2 系統間の D は 0.065±0.0012 で,アキタクロナガオサムシの年代スケールで換算すると 2600±48 万年となる。アルプス隆起は約 2000 万年前に完成したと推定されるので,誤差範囲内で進化距離とほぼ一致する。
進化距離を縦軸に,年代を横軸にして上述のデータを用いてグラフを作成した(図1)。この図からオサムシの ND5 遺伝子の進化速度はほぼ一定と考えてよい)
 |
【図1】
骨片オサムシ群 (Digitulati) の系統関係と日本特産のオオオサムシ亜属の起源
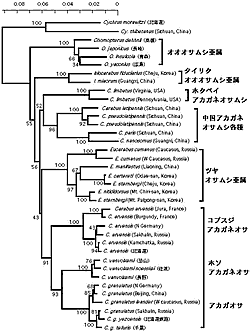
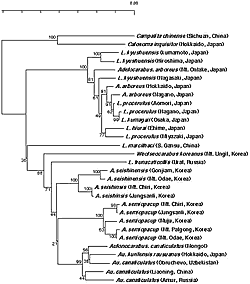
骨片オサムシ亜群は雄交尾器の内袋腹面中央に指状片 digitulus(交尾片または骨片)を備えていることが最大の特徴で、ユーラシア大陸から北米大陸まで、全北区にわたる広い分布域を持っている。井村 (1996) によれば、この亜群はアカガネオサムシ亜属 (Carabus)、ツヤオサムシ亜属 (Eucarabus)、タイリクオオオサムシ亜属 (Isiocarabus)、と日本特産のオオオサムシ亜属 (Ohomopterus) の4 亜属からなるとされる。世界各地から集められたこれらのオサムシのミトコンドリア ND5 遺伝子を解析し、系統樹を作成した。この系統樹によると、骨片オサムシ亜群は少なくとも6つの系統に分かれることが判った(図2)。(1) アカガネオサムシ (Carabus granulatus)/ホソアカガネオサムシ/コブスジアカガネオサムシ; (2) ツヤオサムシ亜属; (3) 北米のヒメホクベイアカガネオサムシ; (4) granulatus 以外の中国産アカガネオサムシ各種; (5) タイリクオオオサムシ亜属; (6) 日本特産のオオオサムシ亜属。ツヤオサムシ亜属とオオオサムシ亜属はそれぞれ単系統としてグループを形成したが、これまでアカガネオサムシ亜属として纏められたオサムシは少なくとも3つ (上記 1, 3, 4)の独立した系統に分かれたことが示唆された。Isiocarabus は中国大陸,朝鮮半島北部,済州島に分布しており,形態的にOhomopterus に近いと考えられていた。しかし、分析した中国南部と韓国済州島のものは互いに近縁で,Ohomopterus とは全く別系統であった。朝鮮半島南部(韓国本土)には,Isiocarabus が分布していないことから済州島のものは中国大陸から直接渡ってきたとも考えられる。そうなると、Ohomopterus の起源を解明するためには朝鮮半島北部のサンプルの分析が必要と思われる。また granulatus はユーラシア大陸の広い地域、サハリン、日本まで分布しているが、分布域全体にわたる多数のサンプルを分析した結果、DNA の差は極めて小さい。これは granulatus の後翅の発達に関係していると考えられる。
 |
【図2】
いわゆるミヤマオサムシ (Oreocarabus) 亜属の多系統性
ミヤマオサムシ (Oreocarabus) 亜属とされるオサムシはヨーロッパアルプスおよび中国大陸中部に分化の中心を持つ。ユーラシア大陸に広く分布し、20 種以上に分類されている。この亜属は現在暫定的に 5 種群(ミヤマオサムシ、アナバネミヤマオサムシ、クロツヤミヤマオサムシ、ウラジミールスキィミヤマオサムシ、ラトローミヤマオサムシの各群)に分けられている。今回、そのうちの 3 種群に属する 9 種、および北京郊外で発見した O. titanus に近縁と思われる新種 O. sui について、ミトコンドリア ND5 遺伝子の塩基配列 (1083 bp) を決定し、日本とフランスに産する代表的なオサムシ亜族の種と共に系統樹を作成して系統関係を検討した。分子系統樹から形態による種群の暫定分類と異なる結果が得られた。ミヤマオサムシ亜属は全体のオサムシ系統樹の中で単系統群にはならず、少なくとも 6 つの独立したグループを形成している。特にミヤマオサムシ種群に分類されている種は、3 つ以上の系統に分散している。また、これらのグループはオサムシ亜族の他のオサムシ亜属の短期間で放散分岐した時とほぼ同時期に分かれていることがわかった。以上の結果から、形態的に1 亜属に分類されてきたOreocarabus はおそらく多系統であることが示唆され、いくつかの独立した亜属に分割した方がいいと思われる。
オーストラリアオサムシとチリオサムシの系統的位置
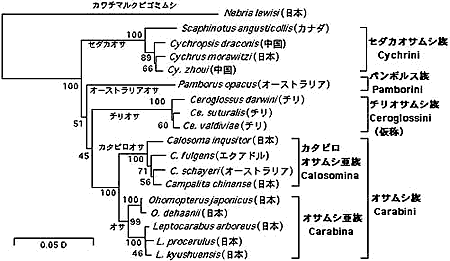
いわゆるオサムシはオサムシ亜科 (Carabinae) に属し,セダカオサムシ族 (Cychrini) とオサムシ族 (Carabini) に分けられ,オサムシ族はさらにカタビロオサムシ亜族 (Calosomina) とオサムシ亜族 (Carabina) に分割されている。mtND5 遺伝子による系統解析の結果は上記の高次分類を支持している。しかし,Carabinae に属するチリオサムシ (Ceroglossus) とオーストラリアオサムシ (Pamborus) の系統分類学的位置については定説がなく,未知である。オーストラリアオサムシについては Kryzhanovsky (1976) がパンボルス族 (Pamborini) を設けているが,Moore (1966) は幼虫の形態からセダカオサムシに近いと主張している。我々の mtDNA と核 28S rRNA 遺伝子による解析では,オーストラリアオサムシ,チリオサムシ,セダカオサムシ族およびオサムシ族はそれぞれ独立の系統となり,ほぼ同時に分岐している (図3)。従って,オーストラリアオサムシとチリオサムシはそれぞれ別族 (Tribe) を形成していると考えられる。
 |
【図3】
カタビロオサムシ亜族 (Calosomina) の一斉放散
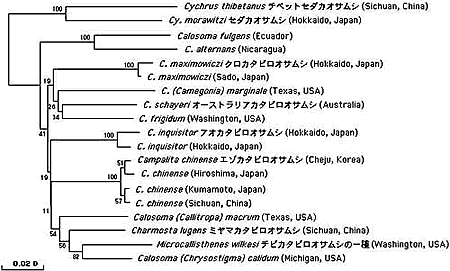
カタビロオサムシ亜族は唯一飛べるオサムシグループで,オーストラリア大陸を含む世界中に分布しており,約150種が知られている。中国,韓国,日本,アメリカ,ニカラグア,エクアドル,オーストラリアなどの国から採集した12種,17個体の mtND5 遺伝子の塩基配列を決定し,系統解析を行った。その結果,これらのオサムシはほぼ同時に分かれ,オサムシ亜族 (Carabina) で見られたような一斉放散がカタビロオサムシ亜族でも起きたことが分かった(図4)。
 |
【図4】
クロナガオサムシ亜属(Leptocarabus)の並行進化
クロナガオサムシ亜属はユーラシア大陸東部から朝鮮半島、日本列島にかけての環日本海地域を中心に分布し、約 20 種が知られている。この亜属は形態的に次の5種群に分けられている。1)クロナガオサムシ種群(中国のクロナガ類と日本のクロナガ類)、2)チョウセンコクロナガオサムシ種群(日本のコクロナガと朝鮮半島のコクロナガ類)、3)チョウセンクロナガオサムシ種群、4)セスジクロナガオサムシ種群、5)チビクロナガオサムシ種群。しかし、mtND5 と核 28S rRNA 遺伝子による系統解析の結果は形態による分類を支持せず、クロナガオサムシ亜属は次の 3 つの系統に分かれた。1)中国のクロナガ、2)日本のクロナガ類/日本のコクロナガ、3)朝鮮半島のクロナガ/朝鮮半島のコクロナガ類/セスジクロナガオサムシ種群/チビクロナガオサムシ種群。つまり、日本のすべての Leptocarabus は共通祖先に由来した単系統である。一方、形態的に極めて近縁とされてきたコクロナガオサムシ種群のもの同志は単系統を形成せず、日本のコクロナガと朝鮮半島のコクロナガはそれぞれ別の系統に属された。このことは、コクロナガオサムシが日本と朝鮮半島で独立に平行して生じたことを示唆している(図5)。
 |
| 【図5】 |
オサムシにおける進化の「動」と「静」
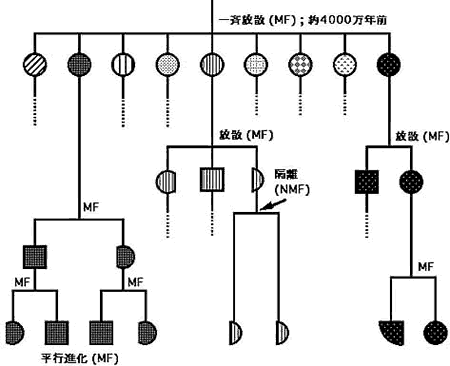
オサムシ亜族 (Carabinae) の主要な(亜)属または群(division)は約4,000万年前に、一斉に放散している。同じような一斉放散はカタビロオサムシ亜族(Calosomina)(上述)やセダカオサムシ族(Cychrini)でもみられる。一斉放散の時期は、インド大陸がユーラシア大陸に衝突し、ヒマラヤ山脈が形成された時期にほぼ一致する。急激な地殻変動に伴う環境変化が、短期間での形態多様化を誘発したと推定している。一斉放散後、それぞれの系統で、中、小規模の放散が起き、現在のオサムシの種が出揃ったと考えられる。中、小規模の放散はヨロイオサムシ群(Procrustimorphi)や骨片オサムシ群(Latitarsi)をはじめとするほとんどすべての群でみられ、形態変化はヨロイオサムシ群でもっとも顕著、骨片オサムシ群ではそれほどでない。さらに、ある形態種から、別の形態種への転換もしばしば起きたと推定される。チベット高原から四川省にかけて分布するチベットオサムシNeoplesius spp.(N)は単系統で、~1,300万年前に一斉に放散しているが、同所的に分布する。頭胸部が著しく肥大したタカネオオズオサムシEocechenus (E)もこの中に入る。N→Eの形態変化が急速に起きたことを示唆している。このような頭胸部の肥大化は、サドマイマイカブリDamaster blaptoides capito や、韓国のオオズクビナガオサムシAcoptolabrus mirabilissimus 、その他でもしばしばみられるが、系統樹ではサドマイマイは本土のコアオマイマイカブリ (D. b. babaianus ) 中に埋没し、オオズクビナガは同所的に産するリーチホソクビナガ(A. leechii )とmt DNAでは全く区別できない。アキタクロナガオサムシEuleptocarabus porrecticollis は、~1,000万年前に本州(多分中部)でマークオサムシLimnocarabus clathratus aquatilis から急激な形態変化を伴って分化したと推定される。これらの例(まだ外に多数ある)は、非連続形態変化が、時には大規模に、またある時には小規模に多様化の過程で起きたことを示唆している。非連続な大変化(explosive evolution)は、動物の門の一斉出現(カンブリアの大爆発)やアフリカのビクトリア湖およびその周辺の湖におけるカワスズメ(cichlids)などが有名で、オサムシの一斉放散に限った話ではない。
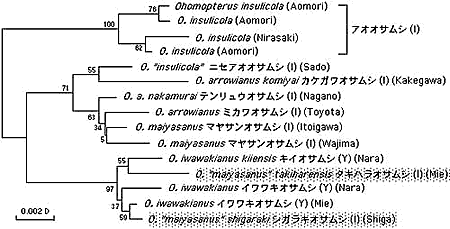
同じような形態変化は時に別系統に並行して起こることがあり、平行進化(parallel evolution)と呼ばれる。日本のオオオサムシ属Ohomopterus は、主として♂交尾器の骨片の形態からヒメオサjaponicus (J)、オオオサdehaanii (D)、ヤコンオサyaconinus (Y)、アオオサinsulicola (I)の4タイプ(種群)に分けられ、それぞれの中には多数の種や亜種が設定されている。mt DNAの系統樹では、形態の区分けのようにはならず、地域的な8系統が認められ、一つの系統に複数のタイプ(たとえばDとJ;IとJのように)が混在する。我々はこの結果を、Ohomopterus の分化の過程で、各地域で同一タイプが独立に平行的に生じたと推定し、これを平行放散進化、その変化の様式をタイプスイッチングと名付けた。
タイプスイッチングの仮説を検証するために、mtDNAの系統樹と核DNAのそれが矛盾しないことを確かめる必要がある。タキハラオサムシ(O. maiyasanus takiharaensis)とシガラキオサムシ(O. m. shigaraki)は近畿南部に局地的に分布し、♂交尾器骨片はIタイプ、一方、同じく近畿南部に広く分布するイワワキオサムシとキイオサムシO. iwawakianus +O. i. kiiensis はYタイプである。mt DNAでも、核DNAのITS領域でも、タキハラとシガラキは、イワワキ(Yタイプ)のクラスターに入り、マヤサン(Iタイプ)とは全く別系統となる(図6)。このことは、Y→Iのタイプスイッチングを強く示唆している。
タイプスイッチングを示唆する他の例としてクロナガオサムシ属Leptocarabus をあげる。Leptocarabus は日本以外では主としてユーラシア大陸北部に広く分布するが、その中で朝鮮半島のセイシンコクロナガL. seishinensis とチョウセンコクロナガL. semiopacus は、日本のコクロナガ(L. arboreus )と形態的に極めて近縁とされる(Sタイプと呼ぶ)。しかし、mt DNAと核28S rDNAの系統解析結果は、Sタイプが別のタイプから大陸と日本で多分タイプスイッチングによって独立に、平行して生じたことを示唆している(上述,図5)。
平行進化は生物界でしばしば見られる現象だが、系統的に離れたもので起きた場合は、形態をよく調べれば平行進化と認識することは容易である。しかし近縁種間で起きた場合には判別できないことがあっても不思議ではない。Ohomopterus やLeptocarabus がその例と考えられる。判別可能な例はオサムシの中でいくつも見出すことが出来る。マークオサ/アキタクロナガオサとトゲオサの♂交尾器の基棘、いろいろなオサムシにおける頭・胸部の巨大化、上翅間室の隆起条化、その他。
形態多様化が非連続的に起こるということは、形態変化がほとんど起きない時期があることを意味している。ザウタートゲオサムシApotomopterus sauteri は、中国と台湾に分布し、ごく軽微な形態差でいくつかの亜種に分けられているが、その分岐は古く、~1,500万年前と計算される。マイマイカブリの近畿以西の3系統は形態的に区別できないがその分岐は~1,300万年前である。ヨーロッパに広く分布するクロツヤオサムシPhricocarabus glabratus は形態的にほとんど区別できないが、mt DNAでみるとアルプスを境に2系統に分れ、その分岐はアルプス形成の~2,000万年前と計算される。同じことがムラサキキンオサムシMegodontus violaceus でもみられる。沿海州のクビナガオサムシ(シュレンククビナガ、ホソクビナガ)は北海道のオオルリオサやオシマルリオサに極めてよく似ているが、分岐は~1,500万年以前である。さらに、チベットオサのNeoplesius wagae とN. markamensis は異所的に分布するが(600 km離れている)互いによく似ていて、後者は前者のせいぜい亜種程度とされていた。 mt DNAでみると分岐は1,200万年前にさかのぼる。非連続な形態変化を「動」とすれば、上にあげたものを「静」と表現することができよう。例示した「静」の時期は、実にオサムシ亜族の歴史の1/2~1/3の長期にわたっている。一般には、地理的隔離が種形成の主因と云われるが、果たしてそうであろうか。上に述べたことは、隔離自身は顕著な形態変化の原因とならないことを示唆している。微小な形態変化は多分核DNAの中立に近い(タンパクのあまり重要でない座のアミノ酸を変えるような)変異によるもので、このような変化が長期に亙って数回起きても形態的にみればせいぜい亜種か、同属の近似種止りなのではなかろうか。「生きた化石」としてあげられる他の生物の例は「静」の時期の長い例にすぎず、例外的なものではない。進化は静と動の組合せによると考えられる(図7)。「動」の変化は核DNAのもっと大規模な変異(いろいろある)によるものと推測している。
 |
|
【図6】 |
 |
【図7】
中国と韓国のオサムシ調査
1998 年6月6日から約 16 日間にわたり、BRH と中国科学院成都分院と共同で, DNA 解析用のオサムシ採集を目的とした第2次中国調査が行われ,また7月3日から17日までの15日間にわたり,BRH 単独(順天大学の白種哲教授と済州道民俗自然史博物館の鄭世湖氏の協力を得た)で韓国オサムシ採集も行われた。中国では,四川省北部から甘粛省南部にかけての地域を調査し,カタビロオサムシ,セダカオサムシを含む合計 350 頭以上(少なくとも15種,約20亜種を含む)のオサムシを採集することができた。一方,韓国の方では,南部の済州島,中部の智異山,北部の五台山の3ヶ所を調査し,カタビロオサムシを含む合計14種,18亜種,850頭以上採集という大成果をあげることができた。これらのオサムシサンプルの DNA 分析は既に完了し,オサムシの進化と起源を理解するための新しい知見が得られた。その結果は本報告中に組入れてある。
今後の研究について
これまでに,井村有希博士,冨永修氏,岡本宗裕博士との協同研究により,世界の約 35 ヶ国から集められた 1000 頭を越えるオサムシを分析した。内訳は約100亜属,種数にして約200種以上である。これらの資料は日本各地を始め海外の多くの同好者の協力によるもので,ここに深謝の意を表したい。
オサムシ研究プロジェクトは1994 年に開始して以来,もう少しで5 年になるところである。研究とは,実に果てがなく,完璧に終わることができないものだが,オサムシプロジェクトで得た教訓は始めたら徹底的 (exhaustive) にやることにより,散発的な解析では到達できない全容が見えてくるということであった。この方針を貫いたからこそ,毎年の報告書に書いたように,オサムシの進化,起源に関して,数々のことが解明され,生物の多様化の原理を探る上でも極めて重要な知見が得られたと考える。来年度は中国、ロシアなどユーラシア大陸に分布しているオサムシを中心にして解析を行い、これまで得られてきた成果をまとめ,総合考察を行い,終止符を打ちたい。