年度別活動報告
年度別活動報告書:2012年度
脊索動物と節足動物の共通祖先を理解する 2−2 カドヘリンの構造と機能の解析
小田広樹(主任研究員) 秋山-小田康子(特別研究員)
佐々木瑞希(奨励研究員) 野田彰子(研究補助員)
逸見なつき(大阪大学大学院生)
はじめに
多細胞動物は、細胞と細胞を連結する微細構造(ジャンクションと呼ぶ)を持ち、ジャンクションを巧みに操ることによって体の形を作り上げる。ジャンクションには何種類かのタイプが知られており、その中でもアドヘレンスジャンクションは、多細胞動物で最も広い系統分布を示すジャンクションで、形態形成に最も密接に関わっていることが知られている8)。アドヘレンスジャンクションの主要な接着分子はクラシカルカドヘリン(以後、カドヘリンと呼ぶ)である。このカドヘリンは隣接した細胞に発現する同種のカドヘリンを認識し、結合する。細胞間隙で起こる、この同種カドヘリン間の結合は、細胞外カドヘリンドメイン(EC)と呼ばれる繰り返し構造を介して達成されているが、カドヘリンのECドメインの数や細胞外領域のドメイン構成には、多細胞動物の系統間で多様性がある。カドヘリンの構造はどのように多様化してきたのか? その際、ジャンクションにおける接着の仕組みは変化したのか? さらに、そのようなジャンクションの変化は形態形成の仕組みに影響を与えなかったのか? 本研究では、これらの疑問に対する答えを探究したいと考えている。
これまでの比較解析では、その構造的多様性がヒトデやクモで共通に見られる祖先状態からの独立的な短縮化で説明できることを見いだしている9) 10) 11)。昆虫においては、祖先的状態のN型カドヘリン(16個のECを持ち、神経組織に局在する)と、短縮化され、派生的状態のE型カドヘリン(7個のECを持ち、上皮組織に局在する)が存在する。脊椎動物とホヤのカドヘリンは昆虫のE型カドヘリンとは起源の異なる5個のECを持つ。このようにカドヘリンは、昆虫に向かった系統、脊椎動物へ向かった系統、そしてその他のいくつかの動物系統で、祖先状態から様々に短縮化してその構造が多様化したことが考えられた。しかしながら、カドヘリン分子の短縮化がその接着機能および動物の形態形成に与えた影響については明らかになっていない。
そこで私たちは、キイロショウジョウバエ (Drosophila melanogaster) のDNカドヘリンとDEカドヘリンをそれぞれ祖先的および派生的なカドヘリンのモデルとして使用し、これらの機能を比較することとした。すなわち、DEカドヘリンをDNカドヘリンで人為的に置換したショウジョウバエを作製することで、カドヘリン分子の短縮化が形態形成に与えた影響を検討することを考えた。今年度は、DNカドヘリンの上皮形成および卵形成における機能について解析を行い、DEカドヘリンと比較した。
結果と考察
DEカドヘリンをDNカドヘリンで人為的に置換した個体の作製
DEカドヘリンの突然変異体はshotgun(shg)として知られている。DEカドヘリン突然変異体にユビキチンプロモーターを用いてGFP融合DNカドヘリンを導入し、人為的にDEカドヘリンをDNカドヘリンに置換したショウジョウバエを作製した。これを用いて、DEカドヘリン接合体ヌルとなった胚にDNカドヘリンを発現させた場合、shgヌルの表現型をどの程度レスキューできるかを調べた。
DNカドヘリンの上皮形成能について
DEカドヘリン接合体ヌルとなった胚では、ステージ15以降で頭部および腹部の上皮形成異常や気管の融合不全、マルピーギ管の形成不全が見られる。これにユビキチン-DNカドヘリン-GFPを導入することで、腹部の上皮形成不全をレスキューした。また、気管の融合およびマルピーギ管も正常に形成された。しかしながら、頭部の一部の上皮は欠損したままであり、頭部の陥入が正常に完了しなかった(図3)。
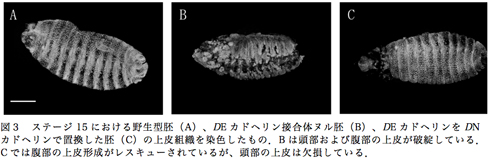
DNカドヘリンの卵形成能について
生殖細胞がDEカドヘリン母性ヌルとなった場合、卵形成に異常が見られる。すなわち、卵細胞の卵室後方に固定されず、正常な位置に局在しない。これに対してDNカドヘリンを導入した場合では、卵細胞は卵室後方に維持された(図4)。

おわりに
今年度の研究により、祖先的な構造状態であると推定されるDNカドヘリンを上皮細胞に発現させた場合、DEカドヘリンと同様に上皮細胞同士を接着する働きを示すことが分かった。また、DEカドヘリンの気管およびマルピーギ管形成における機能についても代替しうることが明らかになった。しかしながら、頭部陥入が不完全であり、頭および顎を形成する能力、あるいはこれらが口陥から引き込まれるという複雑な動きに対応する能力を補うことができないと考えられた。頭部のどの部分に欠損が見られるか、またはどの部位の力により上皮が破綻するのかは更なる追及が必要である。また、DNカドヘリンは卵室内で卵母細胞を正常な位置に保つことができた。今後この卵母細胞から生じうる母性ヌルの胚、つまりDEカドヘリンがDNカドヘリンに完全に置換された胚を、腹溝形成や中腸陥入などアドヘレンスジャンクションの動的制御を必要とする形態形成運動に注目して観察することで、カドヘリンの構造と機能の関係を明らかにしたいと考えている。
引用文献
- 1) Akiyama-Oda, Y., Oda, H. (2010) Cell migration that orients the dorsoventral axis is coordinated with anteroposterior patterning mediated by Hedgehog signaling in the early spider embryo. Development 137, 1263-1273.
- 2) Driever, W., Nüsslein-Volhard, C. (1988) The bicoid protein determines position in the Drosophila embryo in a concentration-dependent manner. Cell 54, 95-104.
- 3) Yazawa, S., Umesono, Y., Hayashi, T., Tarui, H., and Agata, K. (2009) Planarian Hedgehog/Patched establishes anterior-posterior polarity by regulating Wnt signaling. Proc. Natl. Acad. Sci. U.S.A. 106:52, 22329-34.
- 4) Lauter, G., Söll, I., and Hauptmann, G. (2011) Multicolor fluorescent in situhybridization to define abutting and overlapping gene expression in the embryonic zebrafish brain. Neural Development 2011, 6:10 doi:10.1186/1749-8104-6-10.
- 5) Kanayama, M. Akiyama-Oda, Y., Nishimura, O., Tarui, H. Agata, K., and Oda, H. (2011) Travelling and splitting of a wave of hedgehog expression involved in spider-head segmentation. Nature Communications 2, 500.
- 6) Valenta, T., Hausmann, G., and Basler, K. (2012) The many faces and functions of β-catenin. The EMBO Journal 31, 2714-2736.
- 7) Bogdanove, A. J., and Voytas, D. F. (2011) TAL Effectors: Customizable Proteins for DNA Targeting. Science 333, 1843-1846.
- 8) Harris, T. (2012) Adherens Junctions: from Molecular Mechanisms to Tissue Development and Disease, Subcellular Biochemistry 60, Springer Netherlands.
- 9) Oda, H., Tagawa, K., Akiyama-Oda, Y. (2005) Diversification of epithelial adherens junctions with independent reductive changes in cadherin form: identification of potential molecular synapomorphies among bilaterians. Evol. Dev. 7, 376-389.
- 10) Oda, H., and Takeichi, M. (2011) Structural and functional diversity of cadherin at the adherens junction. Journal of Cell Biology 193, 1137-1146.
- 11) Oda, H. (2012) Evolution of the Cadherin-Catenin Complex. In "Adherens Junctions: from Molecular Mechanisms to Tissue Development and Disease" edited by Harris, T., Subcellular Biochemistry 60, pp.9-35, Springer Netherlands.