年度別活動報告
年度別活動報告書:2011年度
脊索動物と節足動物の共通祖先を理解する 2−1 オオヒメグモの胚発生を支配する分子メカニズムの解明
小田広樹(主任研究員) 秋山-小田康子(特別研究員)
佐々木瑞希(奨励研究員) 野田彰子(研究補助員)
西口茂孝(大阪大学大学院生)
はじめに
多細胞動物は形態や発生様式に基づいて動物門と呼ばれる高次分類群に区分されているが、それぞれの分類群を特徴付ける形態が進化してきた過程やその進化の仕組みを理解することは簡単ではない。その大きな要因のひとつは、生物進化において、機能性の高い形態が保存される傾向にある一方で、その形態を生み出すための発生過程やゲノム情報(遺伝子発現制御機構や遺伝子ネットワークなどを含む)は多様に変化しやすいことにある。つまり、形態的特徴によってまとめられた動物分類群の中にも、ゲノムレベルでは大きな多様性が存在しており、もし祖先的状態から大きく変容してしまったゲノムだけで比較解析を進めた場合、進化の歴史を誤って再構成してしまう可能性がある。
例えば、体の軸に沿った繰り返し構造は、ヒトなどの脊椎動物を含む脊索動物門と昆虫やクモを含む節足動物門に共通して見られる形態的特徴である。しかし、両動物門に属する種間で、繰り返しパターンを形作る分子的仕組みに大きな違いがあることを根拠に、この形態的特徴が異なる起源から別々に進化したものであると断定することはできない。なぜなら、それらの動物の祖先において、からだの繰り返しパターンを保持しながらも、そのパターン形成に関わる遺伝子成分や仕組みを大きく変化させた動物集団が存在したかもしれないからである。そういう意味で、それぞれの動物グループの中での多様性を大局的に把握することは重要である。
このような観点に立って、本研究は独自のモデル生物として節足動物門鋏角類オオヒメグモ (Achaearanea tepidariorum) を導入し、既存のモデル生物からの知見と比較することによって、節足動物門の内外での多様性を分析し、ゲノム進化と形態進化を関連づける普遍原理を見出すことを長期目標とする。現段階においては、節足動物門と脊索動物門の間で比較可能な、直交する2つの胚軸(頭尾軸と背腹軸)の形成と頭尾軸に沿った繰り返し構造の形成に着目し、それらの現象に関わる発生プログラムの多様性を調べている。
昨年度までの私たちの研究から、オオヒメグモ胚ではヘッジホッグシグナルが頭尾軸の形成と、背腹軸の向きを決める細胞移動に主要な役割を果たすことが分かった1)。このオオヒメグモ胚での状況は、ショウジョウバエ胚でヘッジホッグシグナルが体節形成の最終段階(体節の極性形成)になるまで必要とされない2)状況とは大きく異なる。ショウジョウバエ胚では、ヘッジホッグシグナルの代わりに転写因子のビコイドが頭尾軸の形成に中心的な役割を果たしている3)。ここで注目すべきは、ビコイド遺伝子がショウジョウバエに近縁の昆虫にしか存在していないことから4)、ショウジョウバエに向かった昆虫系統で頭尾軸形成に関わる遺伝子成分と仕組みに大胆な変化があったことが推定されることである。それに対して、ヘッジホッグシグナル系の成分は多細胞動物の系統に広く分布するため、このシグナル系が異なる動物門の体軸形成を関連づける可能性を秘めている。実際に、扁形動物門のプラナリアではヘッジホッグシグナル系が再生時の頭尾軸極性の形成に関わっていることが示されている5)。
本年度私たちは、ヘッジホッグシグナルに関するこれまでの研究を発展させることによって、遺伝子発現波の分裂を伴う新しいタイプの体節形成をオオヒメグモ胚の頭部領域で見出し、論文発表を行った6)。「結果」としては同じ縞パターンであっても、そのパターン形成を実現させるための過程や仕組みは動物種によって、または体の領域によって様々に異なることが具体的に明らかになった。さらに、初期胚においてヘッジホッグシグナル系の働きを局所的に攪乱することによって、部分的な重複胚を作ることにも成功した。これらの結果は、節足動物門の祖先状態を理解し、この動物門と他の動物門との進化的関係を理解する上で重要な手がかりとなる。
結果と考察
(1)胚盤の予定頭部領域におけるヘッジホッグ発現波の伝播
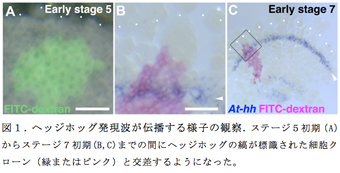
昨年度までの報告書で、ステージ5初期にヘッジホッグの縞状の発現が胚盤の縁に沿って1から2列の細胞で見られ、この後、このヘッジホッグの縞は胚盤の中心方向(後方)に向かって移動することを報告したが、この移動が細胞の運動によるものなのか、遺伝子発現がダイナミックに細胞から細胞へ伝播することによるものかを示す明確な証拠はなかった。今回、蛍光デキストランのマイクロインジェクションによる細胞標識技術を用いて、ヘッジホッグの縞がどのように振る舞っているかを検討した。具体的には、ステージ5初期において、標識された細胞クローンが胚盤の縁から胚盤の中心方向に2細胞以上はなれた位置に存在している胚を、ステージ7初期まで発生させてその標識された細胞クローンとヘッジホッグの縞との位置関係を調べた。その結果、ステージ5初期からステージ7初期までの間に、ヘッジホッグの縞がその細胞クローンの領域と交差するようになったことが分かった。この観察は、予定頭部領域の外胚葉でヘッジホッグの縞が細胞から細胞へ伝播する孤立進行波として振る舞うことを示唆した。
(2)収斂伸長を伴った縞パターンの分裂
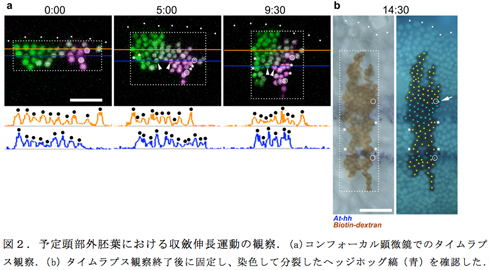
伝播中のヘッジホッグの縞はオルソデンティクルの発現領域の後端と一致しており、両者の遺伝子によって規定される1本の縞は2回の非対称な分裂サイクルを受けて3本のヘッジホッグの縞が生じた。それぞれの分裂サイクルには約4時間かかり、その過程でオルソデンティクル陽性のヘッジホッグの縞が前方に、オルソデンティクル陰性のヘッジホッグの縞が後方に分かれた。細胞クローンでの蛍光タンパクの発現を利用して、ヘッジホッグの縞パターンの分裂が進行している外胚葉における個々の細胞の振る舞いをタイムラプス観察すると、活発な平面内細胞再配列運動による収斂伸長が起こっていることが明らかになった。このことは、ヘッジホッグの縞が分裂する際に生じるヘッジホッグ陰性の領域がヘッジホッグの発現のダイナミックな抑制によって生じることを示唆した。
(3)オオヒメグモ頭部の分裂型体節形成を制御する仕組み
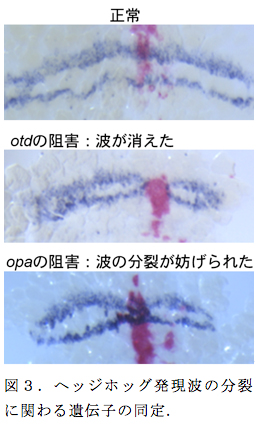
縞パターンの分裂は、これまで数学理論では十分に起こりうることが示されていたが、生物の現象としてはほとんど知られてこなかった。今回、オオヒメグモの頭部領域でそのような現象を発見したことは、新しいパターン形成の仕組みを解明するチャンスをもたらしたと言える。そこで、昨年度に確立した胚性RNA干渉法 (eRNAi) を用いて、予定頭部外胚葉の局所領域で発現を抑制したときに、ヘッジホッグの縞状の発現波の振る舞いに影響が表れる遺伝子を探索した。私たちはこれまでに、ヘッジホッグシグナルによって正に制御される遺伝子として、オルソデンティクルとオッドペアードを同定していたが、これらの遺伝子に対してeRNAiを行ったところ、オルソデンティクルのeRNAiではヘッジホッグの縞が部分的に消失し、オッドペアードのeRNAiではヘッジホッグの縞の分裂が妨げられた(図3)。これらの結果は、オルソデンティクルの活性がヘッジホッグシグナルの正のフィードバックを仲介し、体節形成領域の維持に貢献するとともに、オッドペアードの活性も何らかの形でフィードバック制御に関わって縞分裂のダイナミクスの生成に寄与していることを示唆した。
ショウジョウバエの体節形成では、オルソデンティクルはビコイドの支配下で働く頭部特異的ギャップ様遺伝子として、オッドペアードは空間的周期パターンの形成に中心的な役割を果たすペアールール遺伝子のひとつとして知られている。ショウジョウバエでは上位から下位へ働く遺伝子を順次変えていく階層性の遺伝子ネットワークによって空間的繰り返しパターンが一気に作り上げられるのに対して、オオヒメグモの頭部ではフィードバック機構をもつ遺伝子ネットワークによって時間的に反復される縞分裂が起こり、その結果として空間的繰り返しパターンが生じることが分かった。脊椎動物の中胚葉の分節化では、いわゆる時計遺伝子の活性振動によって空間的繰り返しパターンが生じることが知られているが、今回オオヒメグモの頭部で発見された縞分裂タイプの体節形成は、既存の遺伝子発現の波を起源として「分裂」によって新しい縞を生成している点で「振動」による方法とは異なる(「振動」では既存の波とは離れたところで新しい波のピークが現れることに注意)。根拠は不十分とは言えないが、クモを含め多くの節足動物でも体の後方領域では「振動」による方法で体節形成が行われている可能性が指摘されている。空間的繰り返しパターンを生み出すための方法が様々に存在することは興味深い。これらの現象の比較は対象とするパターンがシンプルであることから、数理モデルによる解析が行いやすく、発生メカニズムの多様化を支配する普遍原理の探究に貢献しうる。この探究は、形態の進化とゲノムの進化の関係をより深く理解することへと繋がると私たちは考えている。
おわりに
本年度は、マイクロインジェクション技術の確立によって可能となったオオヒメグモ胚での細胞標識や胚性RNA干渉を活用して、遺伝子発現のダイナミクスやパターン形成における細胞運動、そして、それらの現象を制御する仕組みを解析し、成果を得た。遺伝子発現と細胞運動を協同させるために仕組まれたゲノム情報は何か。胚発生の調節性を支えるゲノム情報は何か。今後これらの問題に取り組むことは、ゲノムを基盤として動物形態の進化を考えていく上で重要である。また、これらの問題は、数理生物学やシステム生物学とも通じるところがあり、今後そのような方向での取組みも進めていく必要がある。