年度別活動報告
年度別活動報告書:2007年度
脊索動物と節足動物の共通祖先を理解する 2−2 カドヘリンの構造と機能の解析
小田広樹(主任研究員) 秋山-小田康子(奨励研究員)
野田彰子(研究補助員) 春田知洋(大阪大学大学院生)
金山真紀(大阪大学大学院生)
はじめに
多細胞動物を構成する細胞は、細胞間を連結する特殊な微細構造を持ち、それを巧みに操ることによって体の形を作り上げる。このような細胞間結合構造(ジャンクション)には何種類かのタイプが知られているが、アドヘレンスジャンクションは多細胞動物に最も普遍的に見られるジャンクションであり、形態形成に最も密接に関わっている。アドヘレンスジャンクションの主要な接着分子はクラシックカドヘリン(以後、カドヘリンと呼ぶ)である9)。このカドヘリンは隣接の細胞に発現する同種のカドヘリンを認識し、それと結合する。私たちはこれまでの研究で、カドヘリンの細胞外に露出する領域のドメイン構成に左右相称動物の間で多様性があり、その構造的多様性はヒトデやクモで共通に見られる状態からの独立的な短縮化で説明できることを見いだした10)。短縮化されたあるひとつの状態(昆虫型)は、ハエ、コオロギ、トビムシ、アルテミアで見られ、それとは別の状態(脊椎型)が脊椎動物とホヤで、そして、さらに別の状態(頭索型)がナメクジウオで見られる。それぞれの状態は進化的に安定であるが、動物の初期進化では変化できたと考えられる。私たちは、そのようなカドヘリンの構造変化がその分子の機能にどのような変化をもたらしたのかを実験によって検証したいと考えている。その意図は、細胞間接着の機能変化は形態形成に影響を及ぼしうるので、形態進化の仕組みを理解するための手がかりを得ることができると考えたからである。
そこで、ジョウジョウバエの上皮カドヘリン(DEカドヘリン)の構造を一つのモデルとして、カドヘリンの構造と機能の関係を、カドヘリンの突然変異体の表現型をレスキューする実験によって分析した。昨年度までの実験で、無脊索動物のカドヘリンに広く保存されているが、脊椎動物のカドヘリンには存在しない領域を大きく欠く変異型カドヘリン分子(私たちがECC6と命名した分子)が培養細胞において正常分子と同程度の接着活性を示すことを明らかにしていた。このECC6は、人工的に脊椎型カドヘリンに類似したドメイン構成に改変されていると見ることができるが、本年度はこのECC6が個体内の細胞においてどれだけの能力を発揮することができるかを詳細に調べた。
結果と考察
DEEC6はほとんどの上皮組織の形態形成で正常分子と同等の機能を果たすことできるが、頭部の形態形成では十分な機能を果たせない
DEカドヘリンの突然変異体であるショットガン(shotgun,shg)では、(1) 腹部・頭部における上皮の秩序が保持されない、(2) マルピーギ管が細かく分断される、(3) 袋状に陥入したトラキア(気管)原基が隣接する体節間で融合しないなど11)、上皮組織において細胞間接着に関連する致命的な異常が表れる。これらのshgヌル変異体の表現型をDEEC6の発現でレスキューすることができるかを調べた。その結果、腹部上皮、マルピーギ管、気管の表現型はしっかりとレスキューされた。その一方、頭部では脳が露出する異常が観察された。コンフォーカル顕微鏡による頭部上皮のタイムラプス観察から、DEEC6でレスキューを試みた胚は、本来、複雑な細胞運動を行っている頭部上皮に穴が空き、破れてしまっていることが観察された。これは発生初期に脳を形成する神経細胞が陥入したあと、その穴が塞がれないためだと考えられた。
DEEC6は幼虫の上皮組織の発生及び成虫の卵形成において正常に機能できる
shgヌル突然変異体の胚を使ったレスキュー実験では、母親由来の正常なDEカドヘリンの遺伝子産物が卵に供給されているために、胚を構成するそれぞれの細胞においてすべてのDEカドヘリン分子が完全にDEEC6に置き換わっているわけではない。そこで、この問題を回避するためにFLP-FRTを用いた体細胞組み換えによってshg突然変異の細胞クローンを作り、この細胞クローンの表現型がDEEC6の発現によってレスキューされるかを調べた。shgヌル突然変異体の胚を使ったレスキュー実験では、母親由来の正常なDEカドヘリンの遺伝子産物が卵に供給されているために、胚を構成するそれぞれの細胞においてすべてのDEカドヘリン分子が完全にDEEC6に置き換わっているわけではない。そこで、この問題を回避するためにFLP-FRTを用いた体細胞組み換えによってshg突然変異の細胞クローンを作り、この細胞クローンの表現型がDEEC6の発現によってレスキューされるかを調べた。
これまでの文献で記載があるように、幼虫の体内できる翅原基において、shgの細胞クローンは小さなものしか形成されず、大きく成長する前に上皮層から除去されていた。それに対して、DEEC6を発現させたshg細胞は正常に大きなクローンを形成することができた。その細胞の形態や極性関連分子の局在も正常であった。
次に、shgヌルの生殖細胞クローンを作って、卵形成におけるDEEC6の活性を調べた。shgヌルの生殖細胞クローンを持つ卵室では卵母細胞と保育細胞の位置関係に異常が生じる(図3)12)。その卵室では、ボーダー細胞とよばれる体細胞の遊走に異常が表れる。これらのカドヘリン突然変異による卵形成の異常はDEEC6の発現によってほぼ完全にレスキューされた。この結果は、正常なDEカドヘリンを人為的に完全にDEEC6に置き換えた卵及び胚も作出しうることを示唆する。今後は、ドミナントに卵形成異常を引き起こすovoDを組み合わせて体細胞組み換えを行うことによって、正常なDEカドヘリンを完全にDEEC6に置き換えた胚を作製し、その表現型を解析したい。それにより、形態形成におけるDEEC6の分子的能力をより直接的に検証することができると考えている。
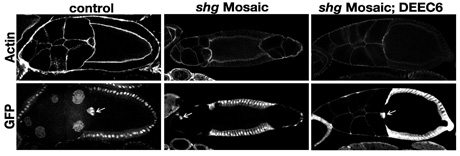 |
図3.卵巣におけるshgヌルの細胞クローンの作出とDEEC6によるレスキュー実験
(左)Control.(中)shgヌル. shgヌル細胞ではGFPの核でのシグナルが観察されない.(右)shgヌルをDEEC6でレスキューしたもの.矢印はボーダー細胞を指している.
おわりに
無脊索動物のカドヘリンに保存されているドメインがなくても、DEカドヘリン分子は生体内でかなりしっかりと機能できることが分かってきた。DEEC6で欠失しているドメインは脊椎動物のカドヘリンには存在しないドメインである。このことは、脊椎動物に向かった系統でカドヘリンが機能を保持したまま大きな構造変化を達成できたことを意味するのかもしれない。しかし、確かにDEEC6は正常分子に近い能力を発揮することができるが、頭部上皮の複雑な形態形成においては十分な機能を果たすことができない。動物進化におけるカドヘリンの構造変化はもしかすると形態の単純化を伴った可能性がある。このような可能性を考慮して、今後もカドヘリンの構造と機能の解析を進めていきたい。
参考文献
1) Carroll, S. B., Grenier, J. K., and Weatherbee, S. D. (2001) From DNA to diversity. Blackwell Science.
2) Holm, A. (1952) Experimentelle Untersuchungen ![]() ber die Entwicklung und Entwicklungsphysiologie des Spinnenembryos. Zool. BiDr Uppsala 29, 293-424.
ber die Entwicklung und Entwicklungsphysiologie des Spinnenembryos. Zool. BiDr Uppsala 29, 293-424.
3) Lane, N. J. (2001) Tight junctions in invertebrates. In: M. Cereijido and J. Anderson (eds.) Tight junctions (second edition), CRC Press LLC, Boca Raton, pp.39-59.
4) Akiyama-Oda, Y. and Oda, H. (2006) Axis specification of the spider embryo: dpp is required for radial-to-axial symmetry transformation and sog for ventral patterning. Development 133, 2347-2357.
5) Oda, H., Nishimura, O., Hirao, Y., Tarui, H., Agata, K. and Akiyama-Oda, Y. (2007) Progressive activation of Delta-Notch signaling from around the blastopore is required to set up a functional caudal lobe in the spider Achaearanea tepidariorum. Development 134, 2195-2205.
6) Oda, H. and Akiyama-Oda, Y. (2008) Differing strategies for forming the arthropod body plan: Lessons from Dpp, Sog and Delta in the fly Drosophila and spider Achaearanea. Development, Growth and Differentiation (in press).
7) Lamar, E., Deblandre, G., Wettstein, D., Gawantka, V., Pollet, N., Niehrs, C. and Kintner, C. (2001) Nrarp is a novel intracellular component of the Notch signaling pathway. Genes Dev. 15, 1885-1899.
8) McMahon, A.P., Ingham, P.W. and Tabin C. (2003) Developmental roles and clinical significance of Hedgehog signaling. Curr. Top. Dev. Biol. 53, 1-114.
9) Takeichi, M. (1995) Morphogenetic roles of classic cadherins. Curr. Opin. Cell Biol.7, 619-627.
10) Oda, H., Tagawa, K. and Akiyama-Oda, Y. (2005) Diversification of epithelial adherens junctions with independent reductive changes in cadherin form: identification of potential molecular synapomorphies among bilaterians. Evol. Dev. 7, 376-389.
11) Uemura, T., Oda, H., Kraut, R., Hayashi, S., Kataoka, Y., and Takeichi, M. (1996) Zygotic Drosophila E-cadherin expression is required for processes of dynamic epithelial cell rearrangement in the Drosophila embryo. Genes Dev. 10, 659-671.
12) Godt, D. and Tepass, U. (1998) Drosophila oocyte localization is mediated by differential cadherin-based adhesion. Nature 395, 387-391.