最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。
バックナンバー
【ミクソゾア:多細胞から単細胞への退化】
2004年12月15日
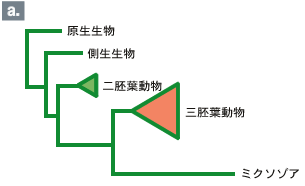
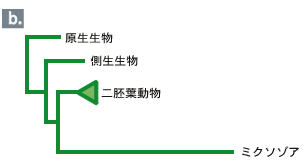
分子系統樹の利点はなんといってもその客観性・再現性および扱いの容易さにある。こうした利点に加え、系統樹推定法が著しく進んだ背景には、方法の改良とともに、データベース、配列解析用ソフト、パーソナルコンピュータ等の普及がある。なかなか分子の結果を信じてもらえなかった時代からすると、現在の普及ぶりは隔世の感がある。しかし分子系統樹法も発展途上にある技術なので、応用に際して間違いを犯しやすい場合が多々ある。ここでは最近の興味深い結果を紹介しながら、分子系統樹法の病理学的側面を考えてみたい。 ミクソゾア類は、魚類や環形動物に寄生する寄生性の生物で、単細胞の真核生物で構成されている原生生物界の一つの門として分類されている。単細胞の生物としてはいささか奇妙な名前が付いている。ゾア(zoa)とは多細胞動物を意味するのだが、それはこの生き物が多細胞の胞子を持っており、その胞子が刺胞動物(二胚葉性の多細胞動物)の刺胞によく似た極嚢を持っていることに由来する。宿主への感染は胞子を経て起こり、極嚢は宿主に付着する際に用いられる。ミクソゾアの極胞が刺胞動物の刺胞に似ていることから、ミクソゾアの二胚葉動物由来が強く示唆されていたが、胞胚が無く、決まった組織を作ることも無いので、仕方なしに原生生物界に放り込んだのだ、とリン・マルグリスは「五つの王国」で述べている。 ミクソゾア門はかつて粘液胞子虫綱と放線胞子虫綱の2つのグループに分類されていた。しかし、粘液胞子虫と放線胞子虫は別々の生物ではなく、同じ一つの生物で世代が違っているだけであることがわかった。今では両者を一つの綱にまとめている。それぞれの世代は異なる生物を宿主としていたので、間違って違う生物だと思ったようである。ともかく1984年の二宿主性生活環の発見は常識を覆す発見として世界中から注目されたようである。 それからちょうど10年後の1994年に、今度は分子系統学から重要な貢献がなされた。18SリボソームRNAを使ってミクソゾアの系統的位置が解明されたのである。それは、なんと三胚葉動物のグループと近縁という衝撃的な結果であった。極嚢から二胚葉性の刺胞動物との近縁関係が示唆されていたが、より複雑な動物のグループと近縁であった(図1a)。これが事実なら、ミクソゾアは三胚葉性の多細胞動物の一系統から、胞子こそ多細胞だが、その細胞は数個しかない単純なもので、むろん神経や消化管など一切ない、単細胞の生物へと体制を著しく退化させたことになる。これは驚くべき進化の出来事といわざるを得ない。実際、この論文に遭遇したとき、「こんなことってあるのだろうか!」と大いに興奮したことを覚えている。※動物の分類については註1を参照※ 翌年には、ミクソゾアの三胚葉性の多細胞動物説に対して強力な助っ人が現れた。ミクソゾアのDNAに多細胞特有の形態形成遺伝子Hox遺伝子が見つかったという論文が報告されたのである。この生物は紛れもなく三胚葉動物由来に違いない、そう多くの人は考えたに違いない。 しかしここに大きな問題が残っている。それはミクソゾアが寄生性の生物であることと関係がある。動物に寄生するためには宿主からの免疫や消化酵素の攻撃など、さまざまな障害をクリアしなければならない。しかし、一度でも寄生に成功すれば十分な栄養が宿主から得られるので、生物の究極の目標である子孫を増やすことに専念できる。そのために感覚器官、運動器官、消化器官など、自由生活する動物には必須な器官が退化してしまう。身軽になった体はさらに生殖能力を高め、逆に退化がいっそう進むことになる。産卵数の増加は生殖細胞(卵と精子)の突然変異率を上昇させ、進化のスピードを高める結果となる(分子進化の中立説によると、進化の速度は突然変異率に比例する)。 進化速度の上昇は分子に基づく系統樹の推定に厄介な問題を引き起こす。分子系統樹では通常、ある生物の系統の枝の長さは進化速度に比例して取られる。従って寄生性の動物では枝の長さが長い。ところで分子系統樹の推定に際し、極端に長い枝同士が結びつけてしまう誤りをしばしば犯す。このことをLong Branch Attraction Artifact (LBA)と呼んでいる。すなわち、寄生性の生物では分子系統樹上の位置を正しく推定することがそもそも難しいのである。 前置きはこのぐらいにして18SリボソームRNAで推定された系統樹(図1a)上でのミクソゾアの位置を見てみよう。確かに主張通り、ミクソゾアは三胚葉性の動物と近縁関係にあるが、2つの枝はいずれも長い。これは上で述べたLBAの可能性がある。つまり、ミクソゾアの系統的位置は正しく推定されていないのかもしれない。 一歩下がって、単細胞のミクソゾアが多細胞動物由来かどうかだけは答えられるかもしれない。それには一方の長い枝の三胚葉動物を除いて推定し直してみるとよい。そうして解析をし直してみると、ミクソゾアは辛うじては二胚葉性の動物に近縁のようである(図1b)。従って、「ミクソゾアは多細胞動物由来で、寄生の結果形態を著しく退化させた」、といった程度の主張なら正しいかもしれない。 では、ミクソゾアからHox遺伝子が見つかっているが、これはどうだろう?この問題は、寄生性の生物から遺伝子を取り出す際の困難と関わりがある。すなわち、宿主の遺伝子とうまく分離できず、いつも宿主遺伝子の汚染(コンタミネーション)に悩まされる。この場合、多数のHox遺伝子を単離しているが、一つを除いて全て宿主の遺伝子か、むしろ宿主の方により似ている遺伝子であった。残りの一つの遺伝子はHox遺伝子そのものというより、Hox遺伝子のコピーで作られた別の遺伝子である可能性が強い。従ってHox遺伝子に関する実験は宿主遺伝子の汚染の可能性があり、これだけでは信頼性がない。 こうしてミクソゾアの系統的位置に関する問題は、寄生動物ゆえの2つの困難に直面して当初の主張を大きく後退せざるを得なくたった。それでも単細胞のミクソゾアが多細胞動物由来である可能性は十分残る。理由は不明だが、リボソームRNAで推定された系統樹では三胚葉動物の系統で進化が極端に速くなっているが、ほかの遺伝子ではそれほど極端ではない。ミクソゾアの正しい系統的位置を知るためにはDNAポリメラーゼやRNAポリメラーゼなどの遺伝子で解析し直す必要があろう。 旧口動物には寄生生活への道を辿ったグループが多くある。中には動物門全部が寄生性であることもある。脱皮することによって異なる環境に適応し易いことと関連があるのかもしれない。寄生性動物を含めて、全ての動物門の系統関係を正確に再現しようとすると、ここで考えた寄生性生物の分子系統樹推定の際に起る2つの困難、すなわち進化速度の上昇に伴う系統樹推定の困難、及び遺伝子単離に際して宿主遺伝子の避け難い汚染の問題は深刻である。この問題の解決は今後の分子系統学に課せられた重要な課題の一つであろう。 |
||||
| ||||
|
[宮田 隆] |