最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。
バックナンバー
【変わる動物の系統樹−三胚葉動物の体腔と系統分類−】
2004年11月15日
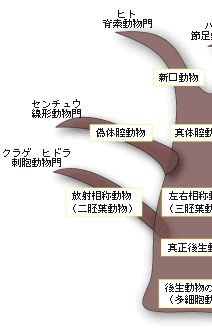
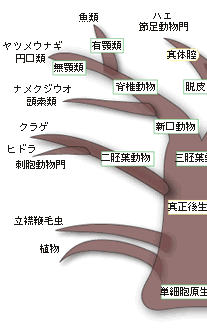
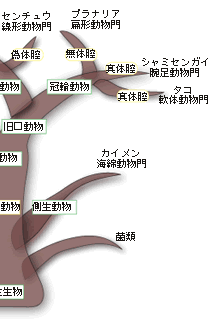
動物は多数の細胞から形作られている多細胞の生物で、植物や菌類とともに一つの生物界を構成している。動物の起源となった生物は単細胞の原生生物で、立襟鞭毛虫のグループが有力視されている。多細胞性の獲得は、ジュリアン・ハックスレーによれば、生物界における生物学的新しさに関する三大革命のひとつで、多細胞化したということは量の新しさを獲得したということで、その結果、体が大きくなり、さまざまな器官や組織を持つ複雑な生物へと進化することができた。大型化に重要な役割を演じたのが体腔、すなわち、細胞層で囲まれ、その中が液体で満たされている空間であった。この空間こそ動物の形態・発生・進化を考える上でたいへん重要な存在なのである。 現存する多細胞の動物のなかで最も原始的な動物はカイメンで、組織もなければ器官もなく、左右の区別もない。細胞の種類は15種類程度で、細胞間の接着や協調などがほとんど見られない。カイメンの細胞はまだ単細胞的で、どの部位でもすべての機能を果たすことができ、必要に応じて別の細胞に変わりうる普遍細胞がいたるところにある。板状動物門のセンモウヒラムシとともに、カイメンは側生動物と呼ばれるグループに分類されている。それ以外のすべての後生動物を真正後生動物と呼んでいる。以下、この類の耳慣れない分類の用語が氾濫するので、図1を参照しながら読み進んでほしい。 |
||||
|
||||
|
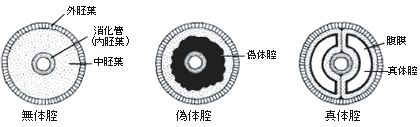
体のデザインの違いから、真正後生動物は放射相称動物と左右相称動物に分類される。放射相称動物は体が放射状に伸びていて、クラゲ、イソギンチャク、ヒドラなど、わずかなグループだけが現存している。ちなみにカイメンのように、対称性のない動物は無相称動物と呼ばれている。放射相称動物は、発生学的には二胚葉性の動物で、左右相称の三胚葉性の動物とは区別される。 放射相称動物や無相称動物を除くすべての多細胞動物は左右相称動物、すなわち三胚葉性の多細胞動物に属する。左右相称動物は、左右が対称で、前後の区別がはっきりしている。体の内部に体腔と呼ばれる空洞があるか無いかで、左右相称動物はさらに無体腔動物、偽体腔動物、真体腔動物の3つに分類されている。 真体腔動物では、内胚葉、中胚葉、外胚葉と呼ばれる3つの異なる組織層が分化し、これら3つの胚葉の細胞集団からさまざまな器官が作られる。中胚葉性組織が広がって体腔を作り、そこに内蔵諸器官が納められることになる。無体腔類にはこうした体腔がない。偽体腔類では空になっている部分はあるものの、真の意味での体腔ではない(図2)。 |
||||
|
||||
|
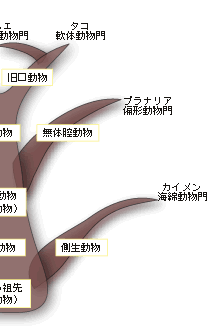
一つの受精卵は発生が進むにつれて、多数の細胞集団からなる、中空のボール状の構造が形成される。ついで、その一端が陥没する。その陥入口を原口と呼ぶ。この原口の運命によって、真体腔動物は、原口が将来の口になるグループ(旧口動物と呼ぶ)と肛門になるグループ(新口動物と呼ぶ)とに分類される。ちなみに、節足動物や軟体動物など多くの動物は旧口動物に属し、われわれヒトを含む脊椎動物やウニなどの棘皮動物は新口動物に属する。 さて、一見、上で述べてきた順に体の複雑さと大型化が増してきているように見える。それははたして進化の順序を示しているのであろうか?たしかに、単細胞の生物から多細胞性を獲得した動物が、上で見てきたような、体の複雑さと大型化を増す方向に枝分かれしながら進化してきたと、一般の教科書には書かれている。この「単純から複雑へ」の進化の傾向は体腔の複雑化の傾向、すなわち無体腔動物→偽体腔動物→真体腔動物の順に一致している。むしろ逆で、体腔の複雑化を基準に再構築された動物の系統関係なのである。まさに体腔は無脊椎動物の比較形態学、系統発生学、系統進化学の要と位置づけられてきたのである。 体腔は動物の体のかなり大きな部分を占め、その中を充満した液体が体中を循環する。このことがさまざまな機能を生む結果となる。体腔内の液体は組織に栄養分や呼吸のためのガスを供給し、細胞からの老廃物の捨て場にもなる。また、体腔はホルモンの循環系としても機能する。なによりも体腔は体の大型化をもたらした。さらに体腔はさまざまな器官を作り、体腔内の液は一種の骨格として働くことで、すばやい運動が可能になった。 こうした体腔が持つ重要な機能を考えると、進化が無体腔動物→偽体腔動物→真体腔動物の順に起きたと考えることは合理的に思える。しかしこの考えは多数派に支持された考えであって、すべての研究者の考えを代表しているわけではない。すなわち、真体腔の体制は動物進化のかなり初期の段階に現れたと考えている研究者のグループがいる。彼らは、無体腔や偽体腔は生活様式の変化に伴って体が小さくなり、その結果体腔に起きた退化的状態だと考えている。教科書的記載はその時代の多数派の意見で、進化研究の最前線ではそれに同意しない、別の意見を持った研究者は何人もいるのがむしろ普通である。 こうした比較形態学や系統発生学における体腔をめぐる意見の対立に対して、分子を用いた系統進化学的研究がアグイナルドらによってなされ、話題になっている。その結果は、これまでの伝統的な系統樹と根本的に異なり、むしろ上記の少数派の解釈に近い。 アグイナルドらはすべての生物が共通に持つリボソームRNAの一つである18SrRNAを使って三胚葉性の動物の系統樹を再現した。その際、彼らはあるトリックを行っている。進化の過程で起こる塩基の置き換えのスピード、すなわち進化の速度が何らかの理由で速い生物の系統は、その系統樹上での位置がしばしば間違って推定される。進化の速度に比例して系統樹の枝の長さを表現するので、この現象をLong Branch Attraction Artifact (LBA) と呼んでいる。このことを避けるために、アグイナルドらは進化速度の速い生物を解析から除外したのである。こうして推定された三胚葉動物の系統樹が図3に示されている。 |
||||
|
||||
|
この系統樹は2つの点で従来の比較形態学や系統発生進化学に基づく伝統的な系統樹と著しく違っている。第一に、新口動物と旧口動物の分岐が三胚葉動物の進化の最初期に起きている(図1では最後)。その後、旧口動物の系統は冠輪動物のグループと脱皮動物のグループに大きく分かれる。これが第二の相違点である。冠輪動物のグループは互いにトロコフォア幼生という共通の形質を持ち、脱皮動物では脱皮が共有形質となる。そして冠輪・脱皮動物のそれぞれのグループには無体腔、偽体腔、真体腔の動物が入り混じっている。すなわち体腔の形態が系統を反映していないのである。第三の相違点は、この系統樹から考えられる自然な解釈は、真体腔の獲得は三胚葉動物の初期進化の過程で起きたということである。従って、無体腔や偽体腔の形態は退化的な形態ということになる。 アグイナルドらの研究は、動物の重要な系統、すなわち動物門の間の系統関係に対して革命的な結果をもたらしたが、18SrRNAのみによる系統樹の推定であり、かつ進化速度の速い系統を意図的に除外した上での解析であるため、十分信頼性のある結果とは言い難い。そもそも現世の動物門はカンブリア爆発によって短期間に成立したと言われているように、厳密な系統関係の推定は難しいのである。現時点では彼らの系統樹は仮説の段階であり、最終結論は今後の研究を待つ必要があろう。多数の遺伝子による、より厳密な系統解析が現在進行中だが、アグイナルドらの系統樹は大筋で正しいように思える。これが事実なら、またしても系統分類に重要な形質を手放しで使うことが出来なくなったことになる。 [宮田 隆] |