

RESEARCH
チョウは脚で味見をする
チョウの味覚から種分化を探る
ナミアゲハはミカン、ギフチョウはカンアオイ…とチョウの幼虫が食べる植物(食草)は決まっています。葉っぱを認識するしくみと葉に含まれる産卵刺激物質との対応を調べれば、チョウの種分化の物語をひもとけるかもしれません。
1.足で感じる植物の味
植物を食べる昆虫の幼虫には、狭食性と言って特定の植物しか食べないものが多い。我々が研究しているアゲハチョウの仲間も、ナミアゲハはミカン科、ジャコウアゲハはウマノスズクサ科と、幼虫が餌とする植物(食草)が狭い範囲に限定されている。これは決して単なる偏食ではなく、長い進化の歴史の中で築かれた仕組みである。卵から孵ったばかりの幼虫は体が小さく、移動能力に欠けるため、自力で餌を見つけることは困難である。そこで、メス成虫による産卵場所の選択が、次世代の生存を左右することになる。メス成虫は産卵する際に飛び回りながら、主に視覚、補助的に嗅覚を利用して幼虫の食草を見つけ出すが、最終的には前脚の先端部の「ふ節」で葉に触れて確かめるのである。ふ節には「化学感覚毛」という器官があり、メス成虫はこの器官で植物に含まれる特定の化合物の組み合わせを認識した上で産卵行動を始める。チョウが植物を見分ける手がかりは、植物の生命維持に直結しない二次代謝物質(註1)であり、植物ごとに組み合わせが異なっている。このような二次代謝物質の中からアゲハチョウの仲間の産卵行動を刺激する化合物(産卵刺激物質)が報告されている。そのすべてが不揮発性なので、「臭い」ではなく「味」として感じていると言っていいだろう。つまりメス成虫は、自分が食べるのではない葉の味見を前脚で行い、幼虫が食べることができる植物に産卵するのである。この食草を認識する仕組みの解明は、まず昆虫と植物を強固に結びつけている絆を明らかにする。しかも種によって食草、つまり認識する化合物が少しずつ変化しているのだから、進化(多様化)における食草転換とその適応的意義を理解する手がかりになるはずである(図1)。

(図1) 食草に産卵するチョウ
[左] ナミアゲハとカラスザンショウ
[中央] キアゲハとイタリアンパセリ
[右]シロオビアゲハとカラスザンショウ
(註1) 二次代謝物質
特に生命維持に関与しない物質のこと。真菌や放線菌がつくる抗生物質なども含まれる。一方、タンパク質、脂質、核酸など、生命活動に必須なものを一次代謝物質と呼ぶ。
2.味見遺伝子の候補探し
昆虫の味覚では、細胞表面にある7回膜貫通型Gタンパク共役型受容体(7TM-GPCR)(註2)が中心的な役割を持つことが知られている。そこで、アゲハチョウの前脚の感覚が味覚の一種であると仮定し、感覚毛で7TM-GPCRが働いているのではないだろうかと考えた。そこで、前脚で働いている遺伝子の情報をできるだけたくさん集めることを目的として、ナミアゲハ雌成虫前脚ふ節のcDNAライブラリー(註3)を作成し、解析した。一万個を超えるcDNAの塩基配列を決定し、コンピュータを駆使して予想されるタンパク質を注意深く調べた結果、前脚ふ節だけで働いている7TM-GPCRが一つ発見できた。仮定から出発した実験の確からしさを一歩進めたことになる。
ショウジョウバエなどから見つかっている味覚と嗅覚の7TM-GPCRのアミノ酸配列を用いて系統解析を行うと、味覚ファミリーと嗅覚ファミリーに分かれるが、我々が発見したナミアゲハの7TM-GPCRは、味覚ファミリーに含まれる(図2)ので、前脚で植物の味見をする際に働いているのだろう。
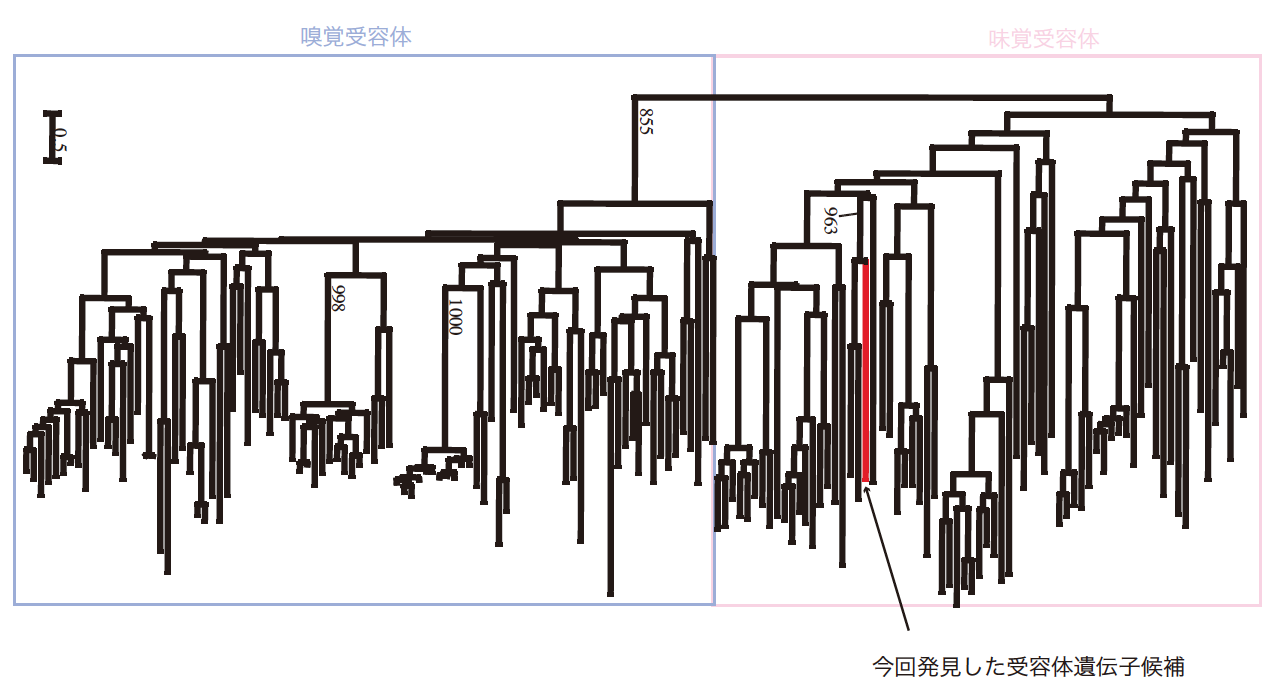
(図2) 嗅覚受容体と味覚受容体の分子系統樹
さまざまな生きものの嗅覚受容体と味覚受容体のDNA塩基配列で分子系統樹を作成したところ、今回みつかった遺伝子は味覚受容体グループに属することがわかった。
(註2) 7回膜貫通型Gタンパク共役型受容体(7TM-GPCR)
Gタンパク質と共益して細胞外の情報を細胞内に伝える受容体。その名の通り、7回膜を貫通する構造をもつ。光や味、匂い、神経伝達物質などの受容体としてはたらく。
(註3) cDNAライブラリー
ある特定の組織や細胞内がつくる全てのmRNAを鋳型にDNAを合成してつくる。スプライシングによってイントロンが取り除かれたmRNAを鋳型とするので、タンパク質に翻訳される情報を知ることができる。
3.本当に味見の遺伝子か確かめる
次に、みつけた遺伝子が産卵刺激物質を認識するために働いているかをどうか明らかにする必要がある。この遺伝子を持っていない培養昆虫細胞にこの遺伝子を持ち込み、ナミアゲハの7TM-GPCRをつくらせて、化合物に対する反応を調べた。発見した7TM-GPCRが産卵刺激物質の受容体であれば、この培養昆虫細胞は産卵刺激物質による刺激に反応し、細胞内のカルシウム濃度が上昇するはずである。カルシウム濃度変化は、カルシウムによって発光するタンパク質(Aequorin:エコーリン)の微弱な光を計測することで測定した(カルシウム・イメージング法)。実は、市販の微弱発光計測装置に独自の工夫を加えて詳細な解析が行えるように改造した結果、この測定が可能になったのである(写真1)。まず、ミカンの葉から抽出した化合物混合液で細胞を刺激したところ、カルシウム濃度が上昇し、7TM-GPCRがミカンの葉に含まれる化合物のどれかに反応することが明らかになった(図3)。次にナミアゲハの産卵刺激物質として知られている10種類の化合物を個別に用いて細胞を刺激したところ、シネフリンとだけがミカン葉抽出物と同じカルシウム濃度の上昇を示した。こうして我々が見つけた7TM-GPCRは、産卵刺激物質のシネフリン受容体であることが分かった。

(写真1)
改良を重ねた手作りの微弱発光計測装置
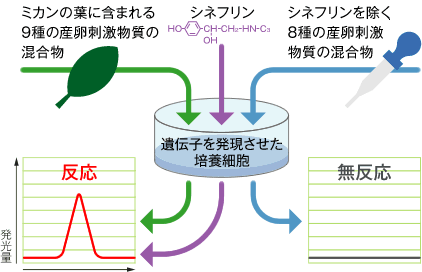
(図3) 何を味見するための遺伝子か調べる
受容体が化合物と結合し、細胞内に信号を伝える際にカルシウム濃度が変化する現象を発光量として捉える装置で測定した。
ミカン科食性のアゲハチョウ3種(ナミアゲハ、クロアゲハ、シロオビアゲハ)で産卵刺激物質として認識している化合物の組み合わせは大きく異なっているが、3種に共通の産卵刺激物質が2種類だけある(図4)。シネフリンはその2種類のうちの一つで、ミカン科食性のアゲハチョウたちにとって重要な化合物だろうと予想できる。
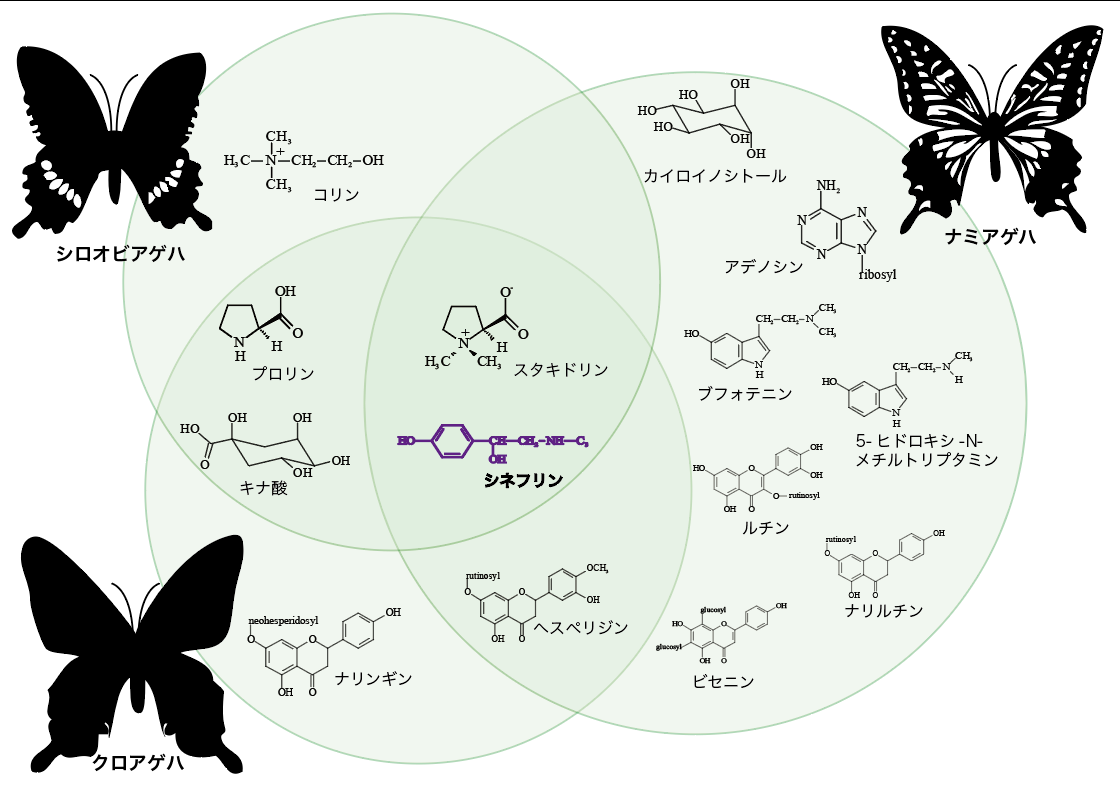
(図4) ミカン科食性のアゲハの産卵刺激物質
シネフリンとスタキドリンが3種のアゲハに共通した産卵刺激物質である。
4.産卵刺激受容システム全体の理解のために
ナミアゲハのcDNAライブラリーから、7TM-GPCRに加えて、化合物を7TM-GPCRまで運搬する役割を持つと考えられる結合タンパク質(CSP)も11種類見いだした。その後、ゲノムライブラリーの解析によりさらに8種類が見つかったので、合計19種類手にした。シロオビアゲハのcDNAライブラリーについても同様の解析を行い、13種類のCSPを見つけた。CSPグループのアミノ酸配列を比較すると、種内(ナミアゲハのCSP同士、シロオビアゲハのCSP同士の比較)では似ておらず、種間(ナミアゲハCSPとシロオビアゲハCSPの比較)ではアミノ酸配列の一致が90%を超える程よく似たものが存在する。これから、CSP遺伝子は種分化以前にゲノム上で遺伝子重複し、それがそれぞれの種内で多様化していったと考えていいだろう。種間で高い類似性を示す相手が見つかった11組について、CSP遺伝子の塩基配列を比較すると、アミノ酸配列が変化しない同義置換の割合はほぼ一定であったのに対し、アミノ酸配列が変化する非同義置換の割合には違いがあった(図5)。このようにCSP遺伝子の保存性のが異なっていることには、何か進化的な意味が隠されているのではないだろうか。これから考えなければならないことである。
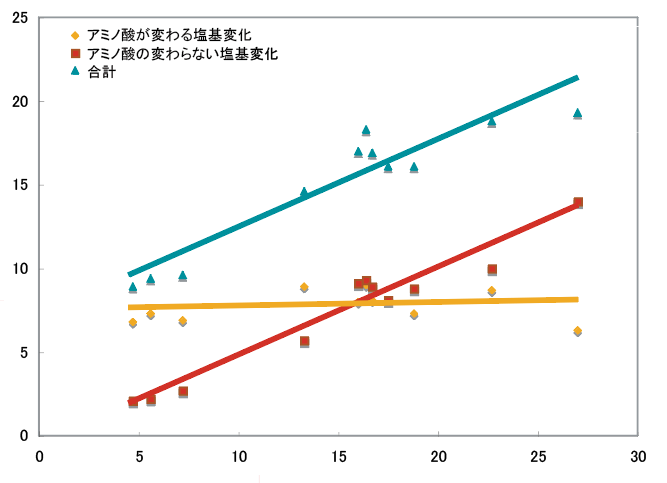
(図5) ナミアゲハとシロオビアゲハのCSP遺伝子の塩基配列変化を比べる
ミアゲハとシロオビアゲハの11のCSP遺伝子の塩基配列をそれぞれ比べると、アミノ酸の配列が変わらない場合(◆)が一定であることが分かった。
5.生命誌の研究らしく
我々の研究は、産卵刺激物質受容システムを構成する分子機構の解明を手がかりとして、昆虫と植物を強固に結びつけている絆や、昆虫たちが子孫を残すために行っている行動の意味を明らかにし、生存の様子をより詳しく理解することを目的としている。また、種内・種間での遺伝子やタンパク質の比較から進化を考えることもできる。このシステムに関わる遺伝子群の一部を発見したことで、本格的な研究に取り組むためのスタートラインに立ったばかりではあるが、生命誌という大きな知に向けての一歩を踏み出せたと考え、さらなる展開を目指している。
尾崎克久(おざき・かつひさ)
2000年弘前大学(岩手連大)博士課程修了。博士(農学)。農林水産省果樹試験場りんご支場生物系特定産業技術研究推進機構ポスドク、JT生命誌研究館奨励研究員を経て2005年より同研究員。
.jpg)

.jpg)
.jpg)










