

RESEARCH
私たちの起源は?
海底に潜むムシから探る脊索動物の起源
ギボシムシという海の砂泥の中で暮らす小さな生きものがヒトへの進化の研究で注目されています。ヒトは脊椎動物ですが、その前を遡るとナメクジウオやホヤと同じ仲間の脊索動物です。「脊索」は、発生過程で背側の正中に現れ、脊椎動物の特徴である中枢神経系(脳や脊髄)をつくるのに重要な器官です。この脊索の起源を探るにはギボシムシに代表される半索動物の研究が重要です。宮本教生さんは、ギボシムシにある「口盲管」に脊索の起源である可能性を見て、中枢神経系の生成の起源を探っています。体づくりの共通性はどこまで遡れるのでしょうか。
1.ヒトの起源をもとめて
我々ヒトの起源はダーウィンによる進化という概念が登場して以来、150年以上にもわたって議論されてきた。ところでヒトの起源と言ってもHomo sapiensの起源を意味する場合もあれば、霊長類に遡ることもある。さらに哺乳類、脊椎動物の起源とより大きな階層を見ることもできる。そしてそれをさらに遡っていくと、われわれヒトとナメクジウオやホヤの属する脊索動物門 (註1)の共通祖先にたどりつくのである。
脊索動物にはその名の通り脊索があり、背側には中空の中枢神経である神経管が走り、鰓裂 (註2)を持つといった特徴がある(図1)。脊索は体の正中を前後軸に沿って走る支持組織で、液胞に富んだ細胞を繊維性コラーゲンの鞘(脊索鞘)が覆う構造をしている。それに加えて脊索は胚発生時にはオーガナイザー(註3)として機能し、その背側に神経管を誘導するのである。この支持組織でもありオーガナイザーでもある脊索と、脊索からの誘導で生じる神経管こそが、脊索動物の最も重要な進化的革新と言える。これらが我々ヒトを含む脊椎動物の脊椎とそこを通る中枢神経である脊髄の形成につながるのである。
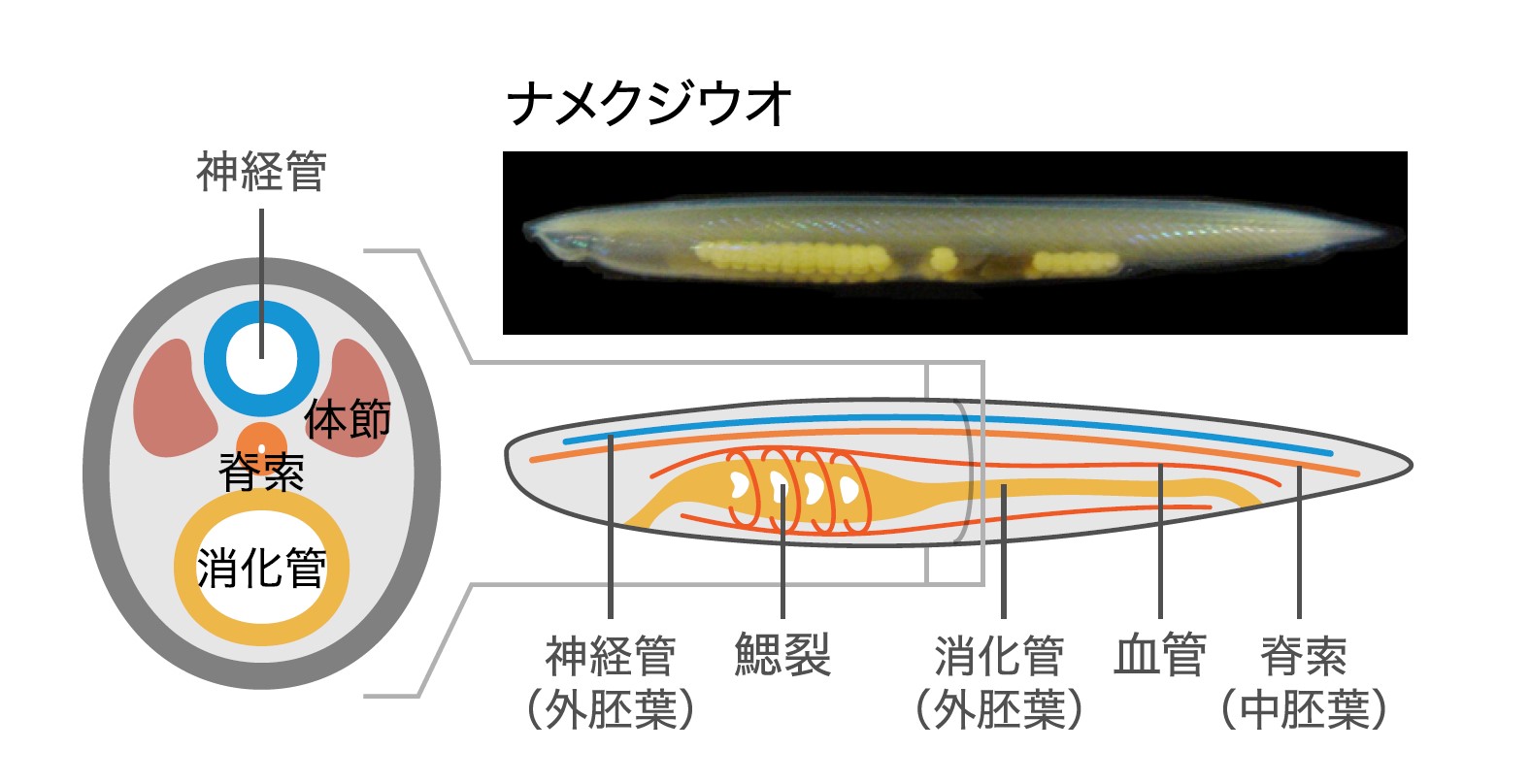
(図1) 脊索動物の体制
最も単純な体制のナメクジウオを例に脊索動物の体制を示す。
19世紀末から20世紀初頭にかけ、脊索動物の体制の起源に関して様々な仮説が提唱されたが、解決の糸口は見つからず、その後しばらくの間議論は停滞していた。しかし近年進化発生学が盛んになるとともに、脊索動物の起源に関する議論が新たな展開を向かえている。
(註1)動物門
動物の分類の階層であり、形態や発生過程に基づいて設定されている。昆虫やエビ、カニなどが属する節足動物門、貝やイカ、タコなどの軟体動物門など約30の動物門がある。
(註2)鰓裂
脊椎動物の胚の咽頭の左右にあり外界に開く孔状の構造。水棲の生きものではここに鰓が生じる。ホヤやナメクジウオなどの脊索動物、ギボシムシでは成体にもある。
(註3) オーガナイザー
胚の発生において接触している領域に働きかけて分化や形態形成を誘導する領域や組織のこと。
2.ギボシムシは脊索動物進化の鍵
脊索動物の起源を知るには脊索動物とその近縁生物との比較が不可欠であり、近縁生物としての第一候補がギボシムシである(図2A)。ギボシムシは長虫状の動物で、海底の砂泥中で丸呑みした砂泥から養分を消化吸収し生活している。現在約100種が知られており、かつては脊索動物の一群とされていた。近年の分子系統解析の結果,半索動物はウニやナマコなどの棘皮動物と姉妹群を形成し、脊索動物はその両方をあわせた仲間の姉妹群という関係である(図3)。棘皮動物であるヒトデなどが五放射相称の体制を持つのに対して、ギボシムシは脊索動物に近い左右相称の体制を持つ。そのことからギボシムシは脊索動物と半索動物・棘皮動物・珍無腸動物(註5)を合わせた後口動物の共通祖先に最も近い形態を残していると考えられている。
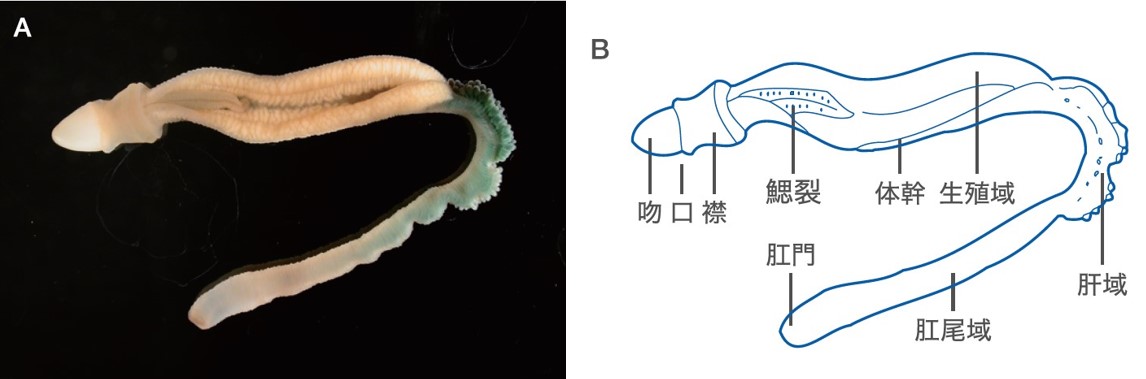
(図2) ギボシムシの構造
ギボシムシは海底に吻で砂を掘って棲んでいる。体長は種類により数センチメートルから最大2メートル程度である。体は軟弱であり採集には注意を払う。
ギボシムシは前方から吻・襟・体幹から構成され、口は吻と襟の間に、肛門は体幹の後端に開いている。また体幹の前方背側には鰓裂があり、これは現生の生物では半索動物と脊索動物にのみ認められる特徴である(図2B)。この鰓裂のほかに、ギボシムシと脊索動物を関連づける非常に重要な形質が2つある。口盲管(stomochord)と襟神経索 (collar cord)である。口盲管は消化管の背側先端が前方(吻の内部)へと陥入した器官で襟部の背側にある(図3)。液胞に富んだ細胞から構成されており、繊維性の結合組織に覆われ、支持組織として機能している。これらの脊索に似た特徴からWilliam Bateson(1886)は口盲管と脊索を相同な器官とみなし、この動物に脊索を半分持つ半索動物(Hemichordata)と名付け脊索動物門の1綱としたのである。口盲管と脊索の相同性に関しては、多くの研究者によって様々な角度から検証がなされ、以下の理由から現在では相同ではないと考える研究者の方が多い。まず口盲管は内胚葉が貫入した構造に過ぎず、中胚葉である脊索とは性質が異なる。また脊索は前後軸に沿って体のほぼ全長に存在するのに対し、口盲管は体のごく限られた領域にしか存在しない。決定的な証拠として脊索動物の発生過程の研究から脊索形成に必須の遺伝子 Brachyury が口盲管の形成時では発現していないことが明らかとなり、脊索と口盲管が相同と考える研究者はほぼいなくなった。
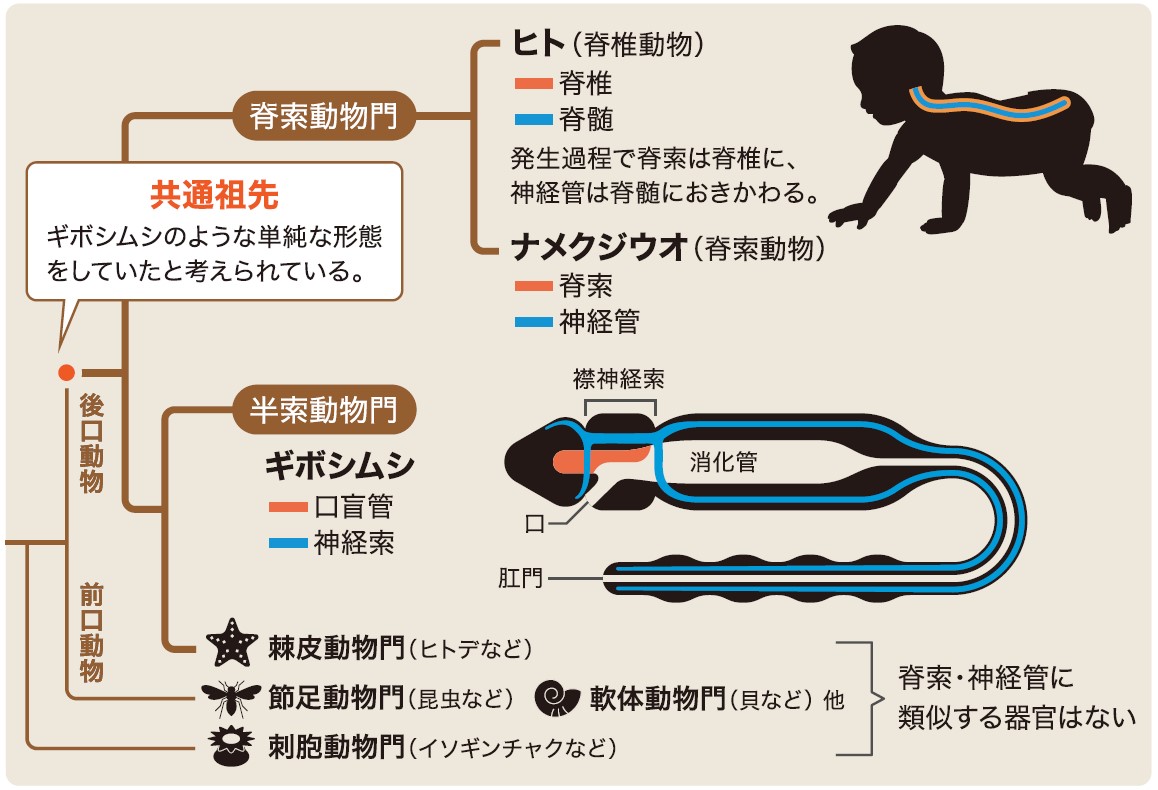
(図3) 動物門における脊索とその類似器官
ギボシムシの属する半索動物門は、ウニやヒトデなどの棘皮動物門と最も近く、その共通祖先で脊索動物と分かれた。脊索と口盲管は共通祖先が持っていた相同器官かどうかが研究の焦点である。
もう一つの形質である襟神経索は、襟部の背側にある管状の神経索であり、脊索動物の神経管と似ている。ギボシムシは背腹正中にそれぞれ1本ずつ、2本の神経索を持っている。背側神経索が吻の基部から襟、体幹へと走っているのに対し、腹側神経索は体幹にのみ存在し、両者は体幹の前端で鰓前神経環によって連絡している(図3)。これらの神経索は基本的に表皮内神経系であるが、襟部でのみ表皮の内側に入り込んでいる。それが襟神経索である。襟神経索の形成過程は、Thomas Hunt Morgan(1894)によって、脊索動物の神経管形成と類似していることが明らかとされ、両者は相同であると考えられた(図4)。ところがギボシムシの背腹を決めるメカニズムの研究の結果,ギボシムシの背腹は昆虫などの前口動物と同じであり、脊索動物とは逆転していることがわかった。これは脊索動物における背腹軸逆転説として形態的にも分子メカニズム的にもよく支持されている。この説によると脊索動物において神経管のある背側は、ギボシムシでは腹側に相当する。つまり背側に存在する襟神経索とは位置関係が矛盾することとなる。襟神経索と神経管の相同性に関しては未だ様々な研究者によって議論され、論争が続いている。
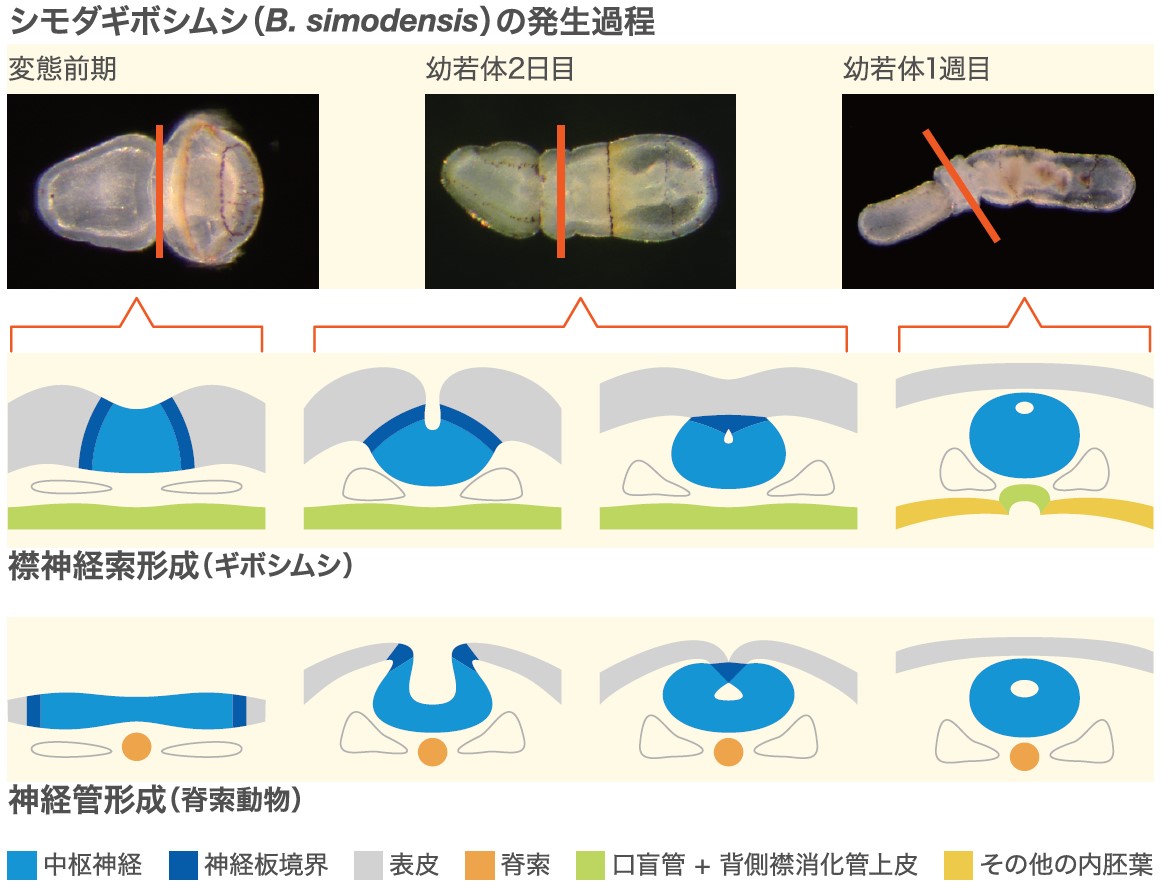
(図4) 神経管形成の比較
Thomas Hunt Morganは、発生過程の比較観察によりギボシムシの襟神経索の形成が脊索動物の神経管形成に似ていることを発見し、これらを相同器官であると提案した。
(註4)神経索
動物の神経系で、同一方向に走る神経繊維の束のこと。
(註5)珍無腸動物
近年新設された後口動物の一群。系統的位置については未だに諸説が存在するが、半索動物と棘皮動物の姉妹群とする説が有力。
3.脊索と神経管の起源と脊索動物の進化
前節で述べてきたように、ギボシムシの口盲管の細胞や組織の形態は脊索に非常によく似ている。脊索との相同性は疑われているものの、実は口盲管がどのようなタイプの細胞から構成されているのか知られていないのである。それはこれまでの研究で用いられていたギボシムシでは、口盲管や襟神経索が形成される変態期の発生過程の観察が困難だったためである。我々は日本にいるギボシムシを改めて調査し、静岡県下田市で発見し,新種として記載したギボシムシ(Balanoglossus simodensis:シモダギボシムシ)の飼育に成功した。こうして初期発生から変態・着底までを観察できるようになったので、口盲管と襟神経索の関係を発生の過程から見直すことにした。そこで脊索の持つ、繊維性コラーゲンを分泌し神経管をオーガナイザーとして機能しているという性質に着目した。脊索で発現している繊維性コラーゲンのギボシムシでの相同遺伝子 ColAの発現を調べたところ、口盲管とその後方の襟背側消化管上皮(前方内胚葉)で発現していた(図5)。またオーガナイザー活性に重要なhedgehog遺伝子(hh)が同じく前方内胚葉で発現していることを確認した。このほかにも前方内胚葉において脊索動物で知られているオーガナイザー関連遺伝子、例えばdkk、 gsc、 otxなどの発現が確認されている。つまりギボシムシの前方内胚葉はオーガナイザーとして機能していると考えられる。
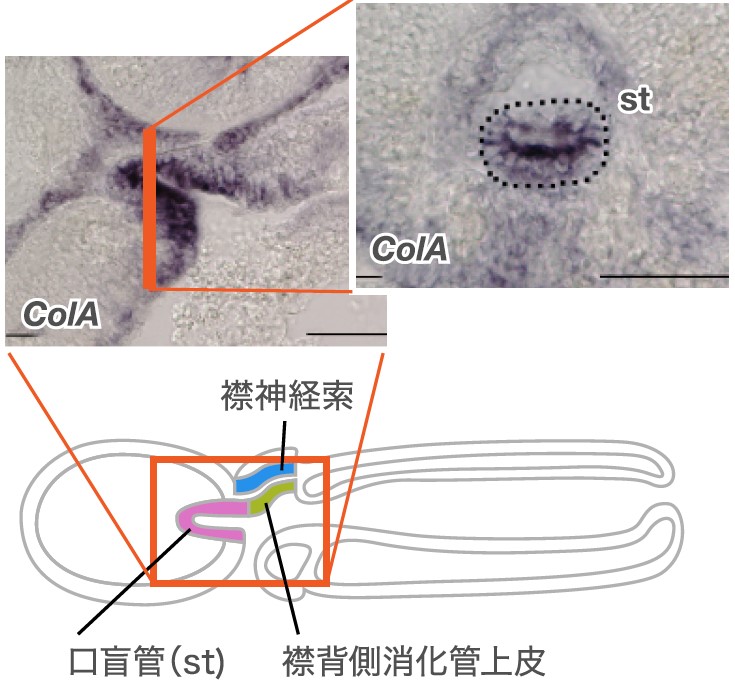
(図5)繊維性コラーゲンのギボシムシ相同遺伝子ColAの発現
脊索の細胞の特徴である繊維性コラーゲンがギボシムシの口盲管にも存在するか調べた。口盲管ができる変態2日目の幼若体では将来口盲管になる領域と隣接する前方内胚葉でコラーゲンの遺伝子(ColA)の発現を確認した。
ギボシムシの襟神経索についても様々なことが明らかとなってきた。襟神経索の発生過程を詳細に観察すると、脊索動物の神経細胞、神経板境界、さらには神経管の領域化に関わる遺伝子が、ギボシムシの襟神経索において、非常に類似したパターンで発現していることが明らかとなった。この結果は襟神経索と神経管の形成過程が、共通のメカニズムを有していることを示唆している。
さらに前方内胚葉と襟神経索は発生の同時期に形成され、前方内胚葉が襟神経索形成に関与している可能性が示唆された。襟神経索は前方内胚葉のすぐ背側で発生し、Hedgehogの受容体をコードする遺伝子patched(ptc)が発現していた(図6)。これは襟神経索の細胞が隣接する前方内胚葉から分泌されるHedgehogを受け取っている可能性を示唆している。このような制御関係は脊索と神経管のそれととてもよく似ている。すなわち前方内胚葉と襟神経索は、それぞれの器官としての類似性だけでなく、その2つの器官の関係性も脊索と神経管の関係と類似していることがわかった。
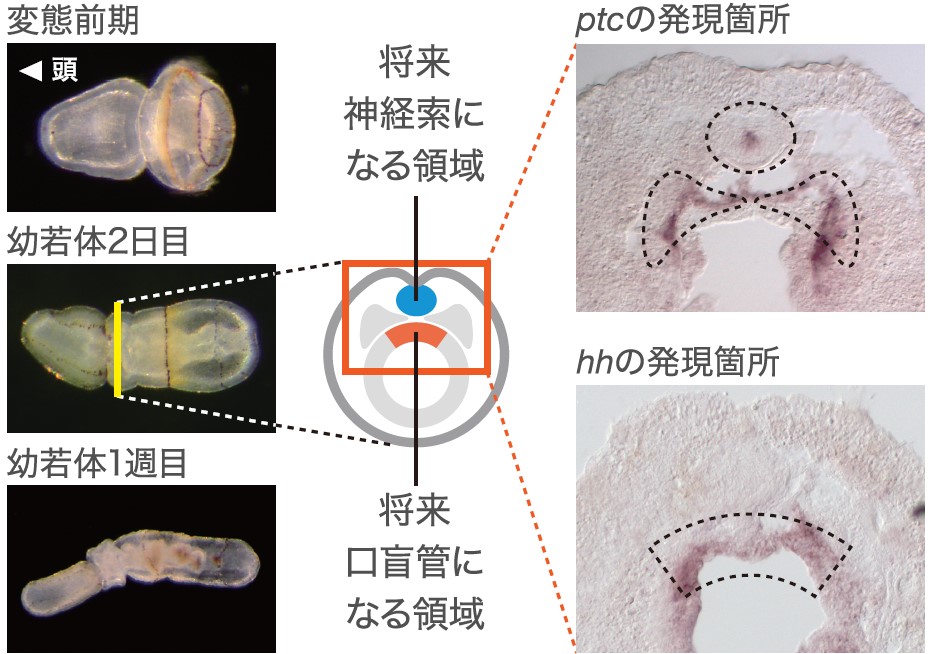
(図6)hedgehog遺伝子(hh)とその受容体patched遺伝子(ptc)の発現
神経誘導のオーガナイザー活性に重要なhedgehogとその受容体のpatchedがそれぞれ将来の口盲管と襟神経索で発現していた。HedgehogのシグナルにPatchedが応答して神経が誘導されていると考えられる。
4.共通のメカニズムから見えてくる後口動物の起源
こうして疑われていた相同性が、発生過程で見られる遺伝子の関係から甦るかのように見えてきた。それでは口盲管と脊索、襟神経索と神経管は、脊椎動物と半索動物の共通祖先にすでに存在していた相同器官なのだろうか。両者が相同と言えればきれいに話がまとまる。しかし、状況は複雑だ。前述の通り口盲管と脊索、襟神経索と神経管は体の中で占める場所が異なっており、発生の由来も異なっているからである。
それを踏まえて現在我々は以下のような仮説を考えている。後口動物の共通祖先において、オーガナイザーからの誘導で管状の神経を形成するメカニズムが獲得された。それがギボシムシでは前方内胚葉と襟神経索であり、脊索動物では脊索と神経管であるというものだ(図7)。どちらが祖先的な状態であるかは現状では判断できないが、少なくともそのような器官を形成する遺伝的な仕組みは共通祖先で既に存在していただろう。両者の違いはその仕組みを発生のどのタイミングで、体のどの場所で使うようになったのかということだ。脊索動物の起源を探るために、今後はメカニズムの共通性をさらに研究する必要がある。例えばギボシムシの前方内胚葉からの誘導による襟神経索形成の仕組みを解明しなければならない。まだ問題はあるものの1世紀以上もの間、仮説が乱立していた脊索動物の起源の問題は、今劇的に解けはじめている。我々ヒトを含む脊索動物の体がどのようにして誕生したのか、その謎が解ける日もそう遠くはない。
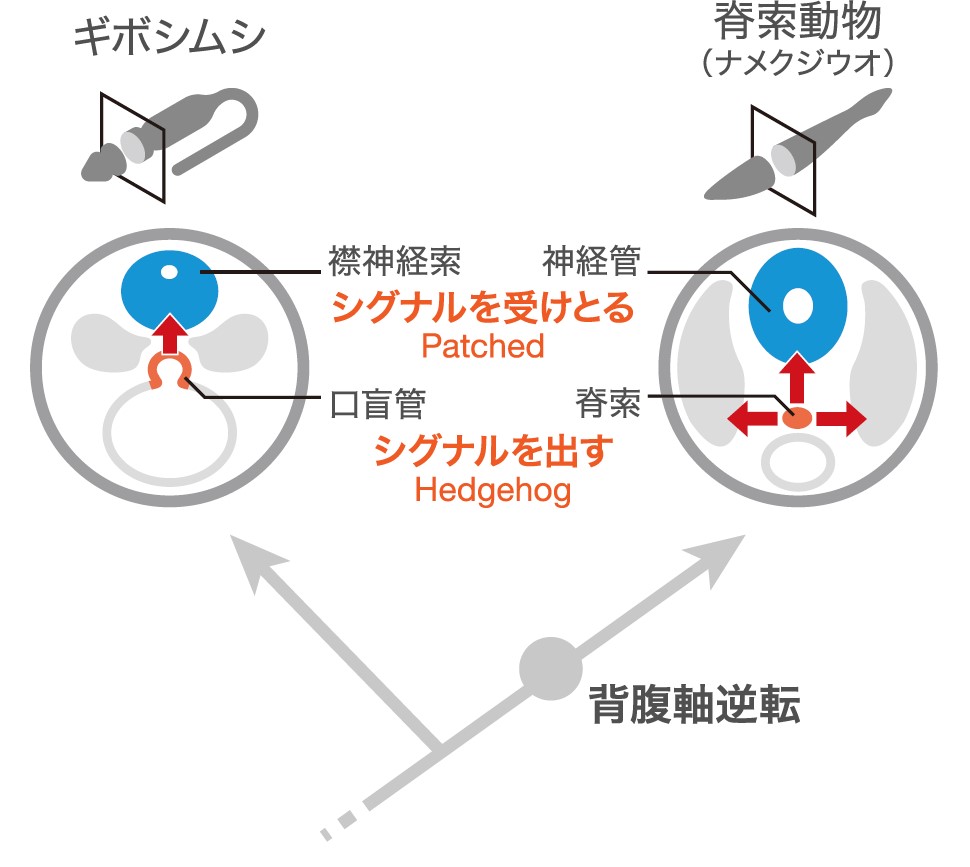
(図7) 口盲管と脊索の関係
口盲管と脊索が相同であるとは言えないが、管状の神経が作られる関係が同一の起源を持っている可能性を示している。

宮本教生(みやもと・のりお)
2011年筑波大学大学院生命環境科学研究科博士後期課程修了。博士(理学)。2011年4月よりJAMSTEC研究員。現在、深海に生息するホネクイハナムシの細菌共生メカニズムや新奇形質「菌根部」の発生メカニズムの研究をおこなっている。
.jpg)

.jpg)
.jpg)










