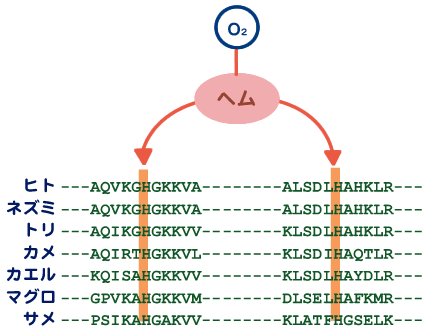
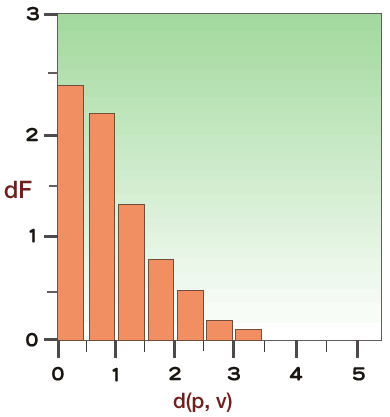
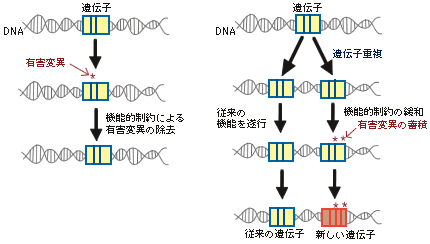
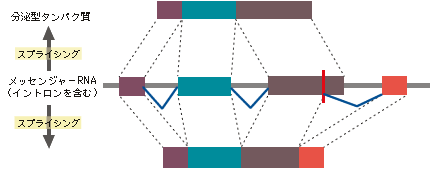
最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。 バックナンバー 【機能シフト:制約からの解放と革新への道】 2005年5月2日 昔、猿の惑星という映画があった。核戦争で人類が壊滅し、それまで抑圧されていた猿が、わずかに生き残った人間を奴隷にして地球を支配するというストーリーである。日の当たらなかったグループがこれまでの支配的グループによる抑圧から解放されることによって、これまでの支配層に代わって大きな展開を示す例は、人間社会に限らず、恐竜絶滅による哺乳類の適応放散など、生物進化における大変革の歴史にしばしばみられる。 こうした支配的生物グループの後退は、残りのグループに対して空っぽの生態的空間(ニッチェ)を準備する結果となり、きわめて多様な形態を持った生物の出現を可能にする。しかし支配層の後退はまれな事件であって、通常は極めて保守性の強い体制下にある。では、こうした保守的体制下にあってどうしたら革新的な道を切り開いていくことができるであろうか。この問題に対して、生物は長い進化の過程で巧妙な方法を編み出し、その方法は多様な生物を創出する上での原動力となった。ここでは、この問題を分子レベルと形態レベルで考えてみよう。分子進化の保守性コピーと組み合わせ肺から鰾への機能シフト 分子進化の保守性 遺伝子やタンパク質が機能する上で重要な部位に突然変異を受け、塩基やアミノ酸が変わると、しばしば重大な機能的変更につながり、最終的に個体の生存に不利に働く。生存に不利な変化は自然選択の力で集団から除去されてしまい、集団に広まることがない。そのためタンパク質や遺伝子が機能する上で重要な部位にあるアミノ酸や塩基は長い進化の過程で不変に保たれることになる。 ヘモグロビン分子を例に分子の保守性を考えてみよう。ヘモグロビンは肺で酸素を吸着し、体の隅々に酸素を運ぶ。さまざまな脊椎動物から取られたヘモグロビンのアミノ酸配列を比べてみると、どの脊椎動物でも常にヒスチジン(H)というアミノ酸で占められている2つの特別な位置(アミノ酸座位)があることがわかる(図1)。すなわち、魚から哺乳類に至る脊椎動物の長い進化の過程で、2つの座位にあるヒスチジンはどちらも不変に保たれている。どちらのヒスチジンも酸素と結合する鉄を空間的にしかるべき位置に固定するのに重要な役割を担っているので、もし別のアミノ酸に置き換えると、鉄の配位を狂わし、本来の機能を損なうことになる。つまりこの座位には変化に対して強い制約が働いているのである。この制約を機能的制約と呼んでいる。 図1. 脊椎動物間でのヘモグロビンのアミノ酸配列の比較アミノ酸配列は一部(ーーー)省略されている。 機能に直接関わる部位以外の領域では、機能的制約は働いていないのか。そうではなく、やはり一定の制約が働いていて、その結果、置き換わっているアミノ酸に一定のパターンがみられる。置き換わるアミノ酸は決してランダムではなく、互いにアミノ酸の性質がよく似ている同士の置換が高い頻度で観察され、似ていないアミノ酸の間では置換頻度が低い。 このことを、アミノ酸の性質をタンパク質の立体構造を決める上で重要な因子である、アミノ酸の体積と極性で代表させることで考えてみよう。こうすることで、2つのアミノ酸の間の物理化学的性質の差を定量的に評価できる。さて、上で定義したアミノ酸間の物理化学的性質の違いの程度と実際のタンパク質で観察されたアミノ酸の置換頻度との関係をみると、両者にはみごとな相関が見られる。物理化学的性質がよく似ているアミノ酸の間では高い頻度でアミノ酸の置換が起きている(図2)。置換するアミノ酸間の物理・化学的性質d(p,v)と置換頻度dFとの良い相関は、アミノ酸の体積や極性といった物理・化学的性質だけで進化の過程で起きたアミノ酸の置換パターンが理解できることを意味している。 図2. 置換するアミノ酸間の物理・化学的性質の違いd(p, v)と置換頻度dFの相関関係 d(p、v)とdFとの間にみられるみごとな相関関係はアミノ酸の変化は許しても、そこには制約があることを表している。タンパク質の立体構造はその固有の機能と密接に関係している。一般に水溶性のタンパク質では、ポリペプチド鎖は球状に折り畳まれて、内部にアミノ酸がコンパクトに詰まった状態になっている。例えばある座位が小さな体積を持つアミノ酸(例えばグリシン)で占められていたとしよう。そのアミノ酸を大きなアミノ酸(例えばトリプトファン)でむりやり置き換えたとすると、タンパク質全体の構造をゆがめる結果になり、最終的には機能に影響することになる。しかし、もしグリシンとあまりサイズの違わないアラニンで置き換えたとすると、タンパク質全体への影響は小さくてすみ、機能も保存される場合が多い。従って後者の変化は前者の変化に比べて有害度がずっと小さく、そのためこうした変化は進化の過程でよく起こるというわけである。これは機能的制約にほかならない。分子の世界では、変化はたとえそれが許されても、本来分子が持っている機能を変えないという制約の下で許される自由度で、きわめて保守的な体制下にあると考えられる。 ▲このページのトップへもどる コピーと組み合わせ タンパク質は、一度獲得した機能を長い進化の過程で保持しつづけるとしたら、どうやって新しい機能を持った遺伝子が進化するのか。実際には生物は進化の過程で非常に多くの遺伝子を作ってきた。生物は細胞内の体制が出来上がった太古の時代から、一つの巧妙な方法で膨大な数の遺伝子を生み出してきた。その方法は、既存の遺伝子の機能を保持したまま、機能的革新をもたらすものでなければならない。生物が採用した方法は遺伝子のコピーを作ることであった。このことを遺伝子重複という。 新しい機能を持った遺伝子を生み出すしくみは次のようなものである(図3)。まず、すでに存在しているある遺伝子の隣にその遺伝子のコピーを作る。こうして出来た一対の遺伝子のうち、一方の遺伝子で従来の機能を果たす。もう一方の遺伝子は役割の上で自由になるので、その遺伝子の上には自由に突然変異を蓄積することができる。一つの遺伝子しかない場合には決して許されない、活性中心や機能的に重要な部位の突然変異であっても、個体に影響しない。なぜなら、もう一方の遺伝子が従来の機能を果たすからである。個体の生存にとって不利な変異を除去する自然選択を一方の遺伝子に集中させ、他方の遺伝子では自然選択の目をそらすことができるわけである。こうして、機能的に重要な部位にも変異を貯めて、新しい機能を持った遺伝子が進化できることになる。遺伝子重複は細胞内の保守的体制下で、革新的な遺伝子を進化させる巧妙な方法で、生物は太古の時代から、もっぱらこの方法で膨大な数の遺伝子を生み出してきた。 図3. 遺伝子の重複と新しい遺伝子の進化 遺伝子重複でいつも新しい遺伝子が進化するとは限らない。コピーの失敗やタンパク質のコード領域に塩基の欠失あるいは挿入が起きて、アミノ酸配列の情報がでたらめになった結果、偽遺伝子(機能を喪失した遺伝子)になることもある。コピーによる新しい遺伝子の生成はまったく試行錯誤の結果なのである。 もう一つ重要な遺伝子多様化の機構が知られている。いくつかの遺伝子あるいはその一部(ドメインと呼ばれる100アミノ酸程度の構造単位)を組み合わせて、一つの大きな遺伝子に統合することで、新しい機能を持った遺伝子が作られる。このことを遺伝子混成(ジーン・シャフリング)という。遺伝子混成の機構によって、わずかな数の遺伝子から多様な遺伝子が作られることは容易に理解できよう。 通常真核生物の遺伝子では、タンパク質を暗号化する部分(エクソン)は連続しておらず、イントロンと呼ばれる情報を持たない塩基配列で分断されている。タンパク質を作る際、まずイントロンをそのまま含んだ前駆体メッセンジャーRNAを作り、次にイントロン部分をはずし、エクソン部分が連結した成熟メッセンジャーRNAに作り変える。このことをスプライシングという。この成熟メッセンジャーRNAがタンパク質合成の際に利用される。スプライシングのパターンを少し変更すことで(オータナティーブ・スプライシングという)エクソンの構成が異なる、違った機能を持つ遺伝子を作ることができる(図4)。つまりオータナティーブ・スプライシングの機構によって一つのDNA領域が機能的に異なる2つの遺伝子を持つことができることになる。この機構は、DNAのサイズに何らかの制約がある場合に多様な遺伝子を作るときにきわめて効果的であろう。これは広い意味での遺伝子混成の一種とみなせるであろう。真核生物は遺伝子重複と遺伝子混成の機構で、驚くほど多様な遺伝子を生み出してきた。コピーと組み合わせこそ、遺伝子多様化の基本的メカニズムなのである。 図4. 一つの遺伝子からオータナティーブスプライシングで 2つの異なるタンパク質を作る。 ▲このページのトップへもどる 肺から鰾への機能シフト よく似た多様化機構が形態レベルにもあり、保守的体制下での器官の機能の変更(シフト)を可能にした例を以下に紹介しよう。動物の細胞は代謝のために絶えず酸素を必要とし、また、酸素の消費に伴って生じる炭酸ガスを体外に放出する必要がある。こうしたガス交換は、水棲の小型無脊椎動物では体の全表面を使って行っている。この方法は、体のサイズの大型化に伴って、早晩行き詰まる。すなわち、体の長さLの増加とともに、体積はLの3乗に比例して大きくなるが、表面積はLの2乗でしか大きくなれないので、結局酸素不足となり、何か別の方法が必要となる。脊椎動物は咽頭由来の鰓という器官を発明することでこの問題を解決した。無顎類のヤツメウナギやメクラウナギにも鰓が認められるので、鰓の獲得は相当古く、脊椎動物の祖先の時代まで遡る。 時代が下がってデボン紀の初期になると、地球が乾燥し古代魚たちが住んでいた川や海が乾きはじめ、水がよどんで鰓呼吸に必要な酸素が不足し始めた。こうした時期に鰓と同じ咽頭由来の器官である肺が硬骨魚類の祖先に導入された。鰓と肺は構造的には非常に違っているが、機能的にはよく似ており、由来も同じであるので、ガス交換用の器官が重複したとみなせるであろう。肺の獲得は初期デボン紀の硬骨魚たちが乾季を生き抜くための適応であったとアルフレッド・ローマーは述べている。 ローマーによると、初期の硬骨魚類は、腎臓の機能からみて、淡水中に生きていたようである。後にその一部の条鰭類のグループが海へと進出していったと考えられている。そこでは海水中に十分な酸素が存在するので、鰓だけで必要量の酸素が獲得でき、結局肺が無用となった。この不必要になった肺が鰾に進化したわけである。まさに無駄が宝を生んだのである。ダーウィンは逆に鰾から肺が進化したと考えたが、この考えは現在では一般に支持されていない。 なぜ鰾という新しい器官が進化できたのかという起源の問題は、自然選択説の困難な問題としてダーウィンの時代からあった。批判の急先鋒はセント・ジョージ・マイヴァートであった。マイヴァート流に鰾の起源に関する批判をしてみよう。いったん鰾が進化してしまえば、より精巧な器官へと進化することは自然選択説で理解できる。しかし、現在の鰾の1%程度の機能しか持たない、出来たての不完全な鰾はそもそも鰾としての役に立つのか。鰾として役に立たないものがどうやって自然選択で進化できるのか。そんなものには自然選択は働かないはずである。もっともな批判である。ダーウィンは種の起源の改訂版で、器官の起源に関するマイヴァートやその他の人々の批判に答えて論じている。ダーウィンのエレガントな解答は、「機能の変化を伴う形質の漸進的推移」であった。鰾は昔から鰾であったのではなく、別の器官から機能を変えたというのだ。 地球の乾燥で鰓呼吸だけでは必要な酸素が補充できないデボン紀初期の魚たちにとって、たとえ現在の肺の1%だけのガス交換の能力しか持たない不完全な肺であっても、その肺を獲得できた魚は生存上有利であったに違いない。自然選択の働きで大きくなった肺が海に進出した段階で不要になり、鰾へと機能を変えることが出来たというわけである。鰓と肺という類似の機能を持った器官が重複していたことが肺から鰾という機能シフトに有効だったことが理解出来よう。 肺・鰾システムは遺伝子重複の機構によって生成された重複遺伝子システムと似ている。いずれの場合もスタートは器官/遺伝子の重複である。また、どちらも一方の器官/遺伝子に強い機能的制約から緩和される時期があり、その緩和を通じて新しい機能を持つ器官/遺伝子へと機能シフトが可能になる。制約が緩和している期間は無駄を抱えることになるので、その期間が長いと無駄な器官/遺伝子はどちらのシステムでも欠失していくことになる。異なる点は、余分な器官/遺伝子が作られるメカニズムの違いであろう。遺伝子の重複は中立な変異に違いないが、鰾の祖先器官を作った変異は、その器官がたとえ不完全であっても、自然選択によって集団に広まった適応的変異であったであろう。それは形態と分子レベルで働く進化機構の違いを表している。 機能シフトには形態レベル並びに分子レベルのいずれにおいても機能的制約の緩和が必須なのかもしれない。器官/遺伝子の重複は制約の緩和を容易にしているようにみえる。重複という道筋を辿らずに同じ器官/遺伝子が進化の過程で機能を変えることがあるだろうか。分子レベルでは、筆者の知る限り、そうした例が知られていない。形態レベルではどうだろう。例えば、クジラ類、海牛類、水生食肉類などの水生の哺乳類はいずれも前肢を胸鰭に変えている。陸上で生活していたこれらの水生哺乳類の祖先はときどき四肢で不器用に泳いでいたのであろうが、水中への進出に伴って歩行と体重を支えるための機能的制約から徐々に緩和され、水かきを持った鰭へと変化していったのであろう。 後肢については興味ある変化がみられる。水生哺乳類では後肢か尾のいずれかを変化させて水中での推進力を作り出す器官を発達させた。アザラシ類、セイウチ類、アシカ類の水生食肉類(鰭脚類)では、陸生の祖先が弱々しい尾しか持っていなかったため、後肢に水かきをつけ、それを後方に折り曲げて推進力を得た。一方クジラ類と海牛類では、尾の先端に水平の尾鰭を持つ独特の構造を進化させた。不要になった後肢は痕跡程度に退化している。四肢の形成にはホックス(Hox)遺伝子が関与するが、Hoxはこれ以外に背骨や生殖器の形成にも関与することが知られている。そのため、後肢は不要になったが、この遺伝子を擬遺伝子化して除去することができない。クジラでは、発生の過程で後肢が不完全な形で形成され、後にアポトーシス(自殺的細胞死)の機構で痕跡程度に小さくしてしまうらしい。これは、形態的には機能的制約が無くなっても、遺伝子レベルでの制約が残っていると、形態が完全に退化することができない例かもしれない。 ▲このページのトップへもどる [宮田 隆] 宮田 隆の進化の話 最新号へ