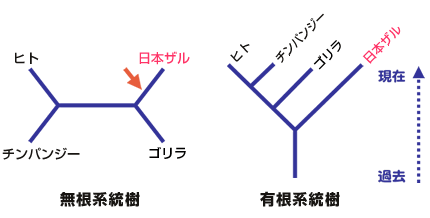
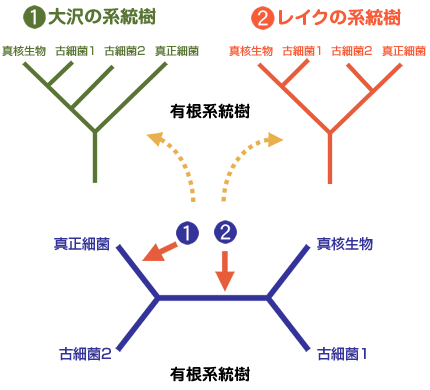
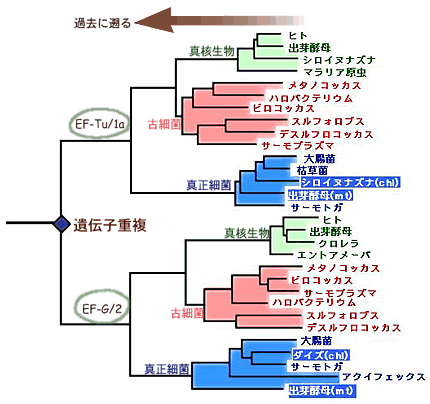
超生物界の間の系統関係が決まると、以後の進化は、外群が決まるので通常の方法で理解できることになる。何はともあれ、誰しもが真っ先に知りたい問題は真核生物の起源の問題であろう。真核生物は古細菌とは独立した系統から進化したのか?あるいは古細菌の一部の系統から進化したのか(側系統という)?そうだとしたらその系統は古細菌のどの系統か?こうした興味ある問題に対して、現時点では確定的な答えは得られていないが、最新のデータからこの重要で興味深い問題について考えてみよう。
最新のNCBI(National Center for Biotechnology Information)の分類によると、古細菌は、ユーリアーケオータ、クレンアーケオータ、ナノアーケオータ、コルアーケオータの4つに分類されている。しかしこの分類は一時的なもので、決して確定的ではない。最近、全ゲノムが決定された超高熱菌を含むナノアーケオータは独自の界を作っているという説があるが、ユーリアーケオータに含まれるという最近のデータもある。現時点では古細菌をユーリアーケオータとクレンアーケオータに分類しておくことには問題なさそうである。
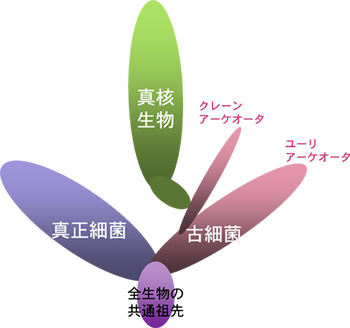
加藤和貴(現、京都大学化学研究所)らは、多数のリボソームタンパクから全生物の系統樹(ユニバーサルツリー)を推定しているが、その系統樹によると、真核生物は古細菌と独立したグループを構成していない。真核生物は古細菌のうちのクレンアーケオータに近縁であり、ユーリアーケオータは両者の外群になっている。この系統関係は、統計的に十分信頼のおける結果になっているが、解析に含まれている系統の数は必ずしも十分ではないので、まだ検討の余地を残している。この結果を信用すると、真核生物は古細菌の一部、すなわち、クレンアーケオータから派生したことになる(図4)。クレンアーケオータから枝分かれした真核生物の祖先細胞は、真正細菌、古細菌のさまざまな系統から遺伝子を水平的に取り込み、真核生物へと進化したのであろう。真核細胞内にみられるさまざまな細胞内小器官と遺伝子水平移動との関連は今後明らかにされるべき重要な問題であろう。