

RESEARCH
「語る科学」
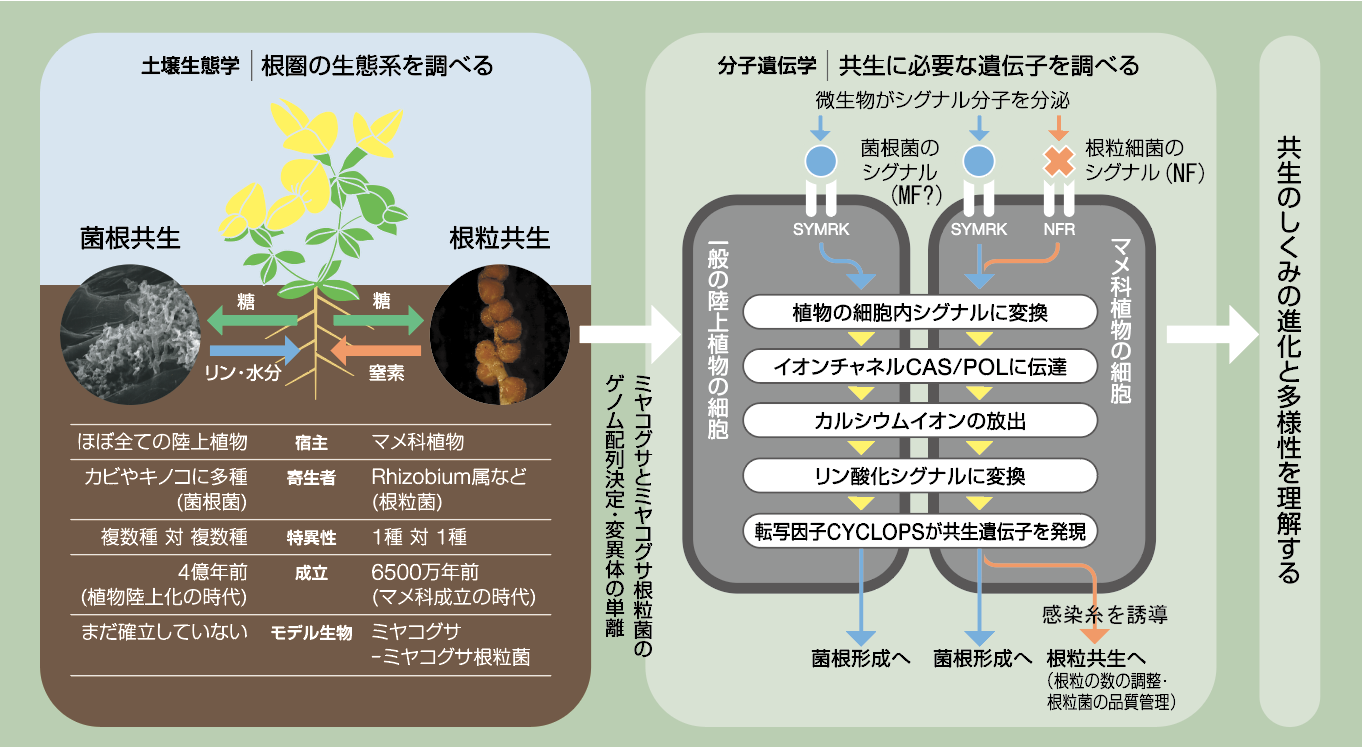
共生のしくみ
-植物と土壌微生物の遺伝子ネットワーク
研究マップ:最新のリサーチを、研究全体の中で位置づけよう。
1.マメの根粒
あたかも春の訪れを謳歌するように野原で花咲くシロツメクサやカラスノエンドウ、秋の気配とともに目に留まるヌスビトハギの節果など、身近なマメ科植物は少なくない。それらの根を引き抜いてみよう。小さなこぶがいくつも付着している。土壌バクテリアの一種、根粒菌が植物体に共生した組織、根粒である【図1】。植物はこの「根粒共生」によって大気中の窒素を利用できるようになる。根粒菌はほぼ例外なく、マメ科植物としか共生しない。
土壌中には多くの微生物が存在し、その多くは植物の役に立たないばかりか害を及ぼす病原菌もある。その中で、マメ科植物は有用な微生物(ゲスト)を選別し、ダイズにはダイズ根粒菌が、アルファルファにはアルファルファ根粒菌が感染するという1対1の関係(宿主特異性)を築いてきたのである。その経緯を知りたい。これが研究の始まりである。

(図1) 植物の根と根粒
根粒共生を行うマメ科植物は、やせた土地で生育できる。ミヤコグサ(Lotus japonicus)は、日本に自生するマメ科の多年草。ミヤコグサとそれに共生するミヤコグサ根粒菌(Mesorhizobium loti)とは共に、ゲノム解析が進んでいる。根粒菌ゲノムの解読は2000年に終了。
2.根毛の有効利用:感染糸
「根粒共生」が始まるには、まず植物がゲストの根粒菌を認識しなければならない。ここに巧みな遺伝子ネットワークのはたらいていることが、近年明らかになってきた。まずマメ科植物の根が分泌したフラボノイドを土壌中の根粒菌が受けとると、根粒菌はシグナル分子ノッドファクターを合成し始める。植物はノッドファクター分子の構造によってその種類を認識し、共生相手の根粒菌だけを根毛に包み込み(カーリング)、筒状の通り道(感染糸)から植物細胞内に取り込む【図2】。根毛は本来、根の表面積を増やして養分や水を効率よく吸収するものだが、マメ科植物では根粒菌の取り込みに利用されている。
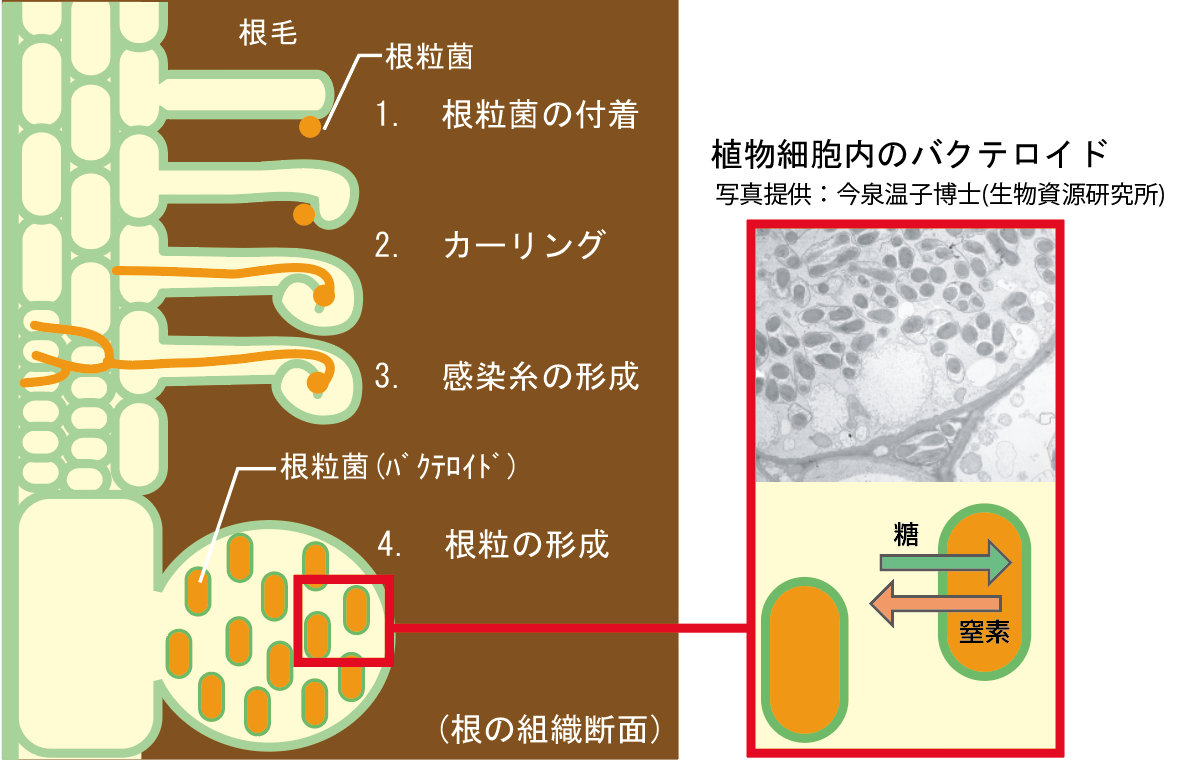
(図2)根粒形成のしくみ
(1.)根粒菌が植物の根毛に付着すると、(2.)根毛は変形して根粒菌を囲い込み、(3.)筒状の感染糸を通して根粒菌を内部に送る。(4.)根の細胞が分裂してこぶ状の根粒をつくり、その中で根粒菌は盛んに「窒素固定」を行う。(5.)根粒菌は大気中の窒素をアンモニアに変換した後に植物に提供し、自身のエネルギー源に植物の光合成産物を利用する。
その秘密は、根毛の表面とつながった感染糸ができること。感染糸の内側は細胞の外になっているので、根粒菌は植物の細胞質と接することなく感染を進行させる。他の植物にこのような例は無い。そして、いったん感染した根粒菌に対して選択を行い、例えば菌表層の多糖類に異常があると、それ以上の侵入を防ぐ。われわれはミヤコグサという、日本に自生する小さな野生マメ科植物をモデルにして、感染糸形成にはたらく遺伝子を調べた【図1】。共生関係にある根粒菌が付着しても感染糸の伸長が途中で止まってしまう変異系統をいくつも見出し、その原因は2つのいずれかの遺伝子座の欠損によることを突き止めた。これらの遺伝子は、根毛内で根粒菌に対し選択を行う際にはたらき、さらに根粒の発達にも影響をおよぼす【図3 -B】。実は、通常の根粒形成の際、感染糸の8割以上は根毛内で止まってしまう。まずは多く感染させておいて、必要な分だけをさらに根粒まで誘導するしくみがあるのだろう。細胞内に入れるのは慎重にという姿勢が見える。
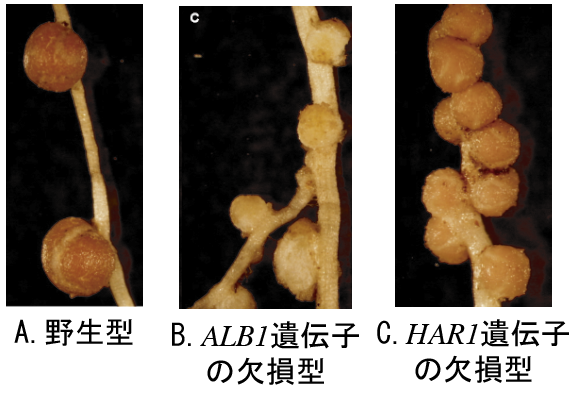
(図3) ミヤコグサ根粒形成の変異体
遺伝子ALB1の欠損したミヤコグサ変異体では根粒が成熟せず、遺伝子HAR1が欠損したミヤコグサ変異体は、過剰の根粒をつくる。
3.外の窒素と内の窒素:根粒の数のコントロール
マメ科植物と根粒菌の共生に見られるもう1つの巧みなしくみは、根粒の数のコントロールである。必要充分な根粒ができると、それ以上は根粒菌を感染させない。根粒をつくりすぎて植物の生長が強く抑えられてしまうミヤコグサの変異体では、遺伝子HAR1が欠損していることが分かった(東京大学の川口博士らの研究)【図3-C】。HAR1は地上部ではたらく受容体タンパク質であり、地下部からの「充分な数の根粒ができた」というシグナルを受け取ると、「根粒を作らせない」という指令を地下部に送り返すらしい。根粒形成の最初のシグナルは、根粒菌のノッドファクターだが、こうしてできた根粒がつくる窒素化合物の量が、第二のシグナルとして大きな役割を果たしているらしい。実は、土壌中に硝酸など根から吸収できる窒素源が充分にあると、マメ科植物は根粒をつくらない。興味深いことに、遺伝子HAR1の欠損型では、土壌の窒素源がある時の根粒数のコントロールも効かなくなっていた。つまり根の外の窒素(土壌中の硝酸)も内の窒素(根粒由来の窒素化合物)も共通のしくみで根粒形成をコントロールしているのだ。
そこで窒素化合物の応答をより詳細に知るために、HAR1以外の因子を調べた。2万個の遺伝子を同時に解析できるDNAアレイで調べたところ、「ジャスモン酸」が浮かび上がってきた。植物は、害虫に葉を喰われるなどのストレスを受けると、ジャスモン酸を全身に循環させ、害虫に抵抗するための遺伝子を発現誘導すると言われている。ところが根粒共生の場合、ジャスモン酸は根粒菌の感染を防御するのではなく、促進するという意外な役割をしている。ジャスモン酸応答遺伝子は「外の窒素」の余剰でも「内の窒素」の余剰でも抑えられ、さらにミヤコグサにジャスモン酸を与えると感染が促進された。ジャスモン酸は、地下部→地上部→地下部と行き来して根粒の数を調節する重要な因子の1つなのだろう。マメ科植物は、植物のストレス応答という普遍的なしくみを根粒共生のために借用し、しかも有用な菌の感染を促進させているというのは興味深い。
4.もう1つの地下共生:菌根
植物と微生物の土壌生態系に、根粒共生より一般的な「菌根共生」がある。こちらは植物にリンや水分、微量元素を提供する。菌根菌はカビの一種で、特にアーバスキュラー菌根菌は8割以上の陸上植物種と共生する内生菌根菌である(他にも、例えばマツタケのような外生菌根菌もある)。根粒菌と異なり、内生菌根菌はホストをあまり選ばず、また単独では培養できない。「菌根共生」では、菌根菌が根の内部に菌糸を伸ばし樹枝状体を展開する過程は知られているが【図4】、植物と微生物の間の認識に関わる遺伝子ネットワークの実態は、根粒菌の場合と違い、全くわかっていないと言ってよい。
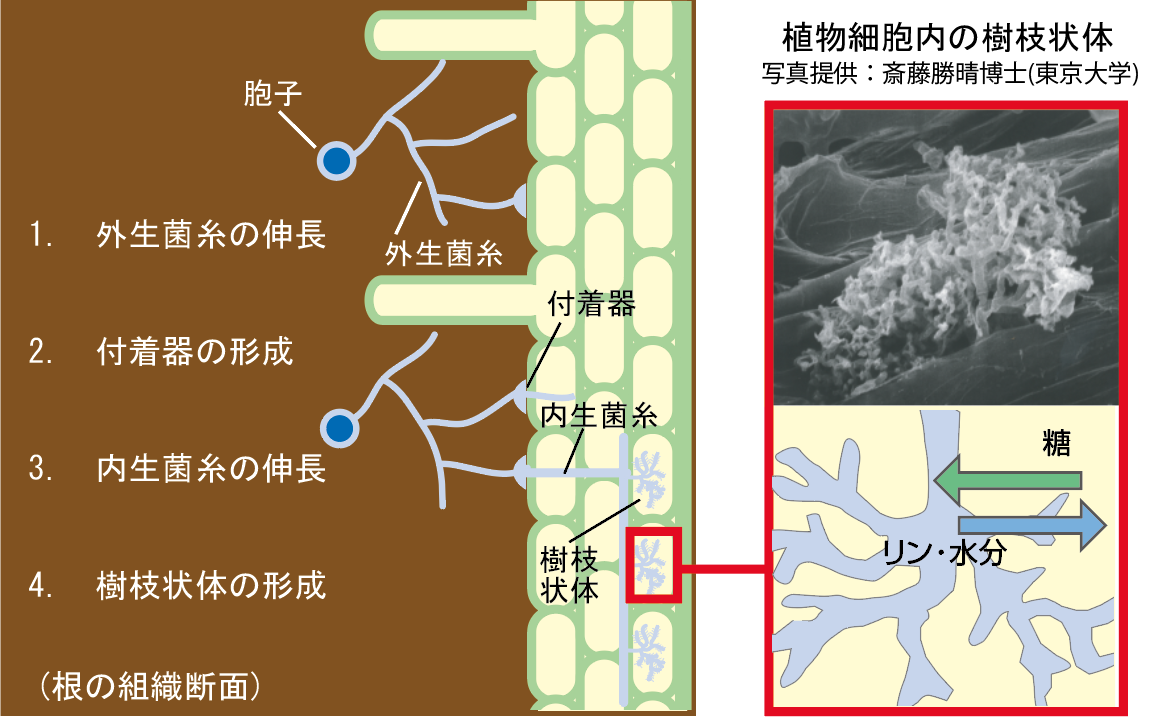
(図4)菌根共生のしくみ(内生菌根菌)
(1.)菌根菌が根を認識すると、(2.)根の表面に付着器を形成し、(3.)そこから内生菌糸を根の内部に伸ばし、(4.)植物の細胞内に樹枝状体とよばれる構造をつくる。(5.) 菌根菌は土壌から集めたリンや水分などを植物に提供する一方、植物の光合成産物を自身のエネルギー源に利用する。
菌根菌は多核なので遺伝的解析が難しく、根粒菌のノッドファクターにあたる最初のシグナルも同定されていない。しかし、幸いにも植物側の遺伝子については解析が進みつつある。実は、ミヤコグサで根粒共生に関わる遺伝子を調べているうちに、根粒形成初期の一連の応答が変異する系統では、同時に菌根共生も破綻するという予想外の事実が明らかになったのである(註1)。これは、根粒共生と菌根共生ではたらく遺伝子はそのネットワークの一部を共有しているということを意味する。われわれは2つの共生が共に不全となる変異系統を解析し、このネットワークに関わる2つのタンパク質CASTORとPOLLUX(CAS/POL)を同定した。これらはプラスチド(細胞内小器官の1種)に局在するイオンゲート型イオンチャネルで、細胞内のカルシウムイオンが周期的に増減する現象(カルシウムスパイキング)に関与していることがわかった。根粒形成初期に植物の細胞内でカルシウムスパイキングが生じるという現象は、10年近くも前に発見されたが、そのしくみは長い間謎とされてきた。CAS/POLが透過するイオンの種類やゲートに必要なイオンの種類(カルシウムで調節されるカルシウムチャネルと仮定している)など、未解決の課題はあるが、このようにして2つの共生にとって重要な鍵となる遺伝子を明らかにし、課題を解いていく道はひらけたと言えよう。
これまでの研究で分かった共生遺伝子のネットワークを図にまとめた。2つの共生に共通する遺伝子の相互作用、つまりシグナル伝達経路、をコモンパスウエイ(共通経路)と呼ぶ。
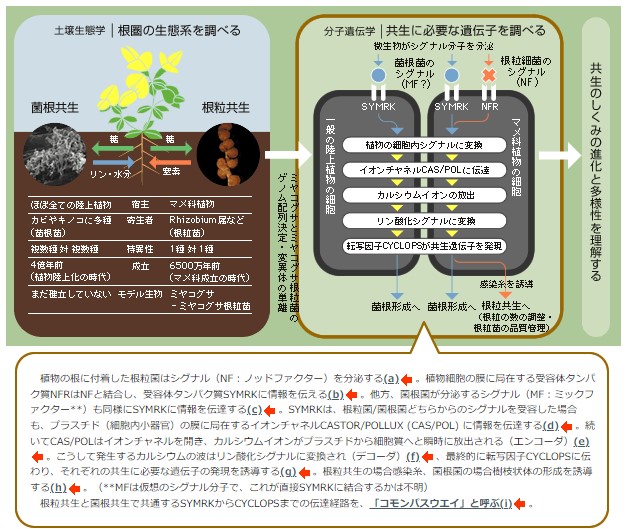
ところで、コモンパスウエイを経由した情報の行き着く先には、代謝やゲストの維持など共生に必要な多くの遺伝子の発現を効率よくコントロールするため、遺伝子のスイッチを一斉に「オン」にする、共生特異的な転写因子が欠かせないであろう。われわれはこの転写因子の候補としてCYCLOPSを同定した。ところで、CYCLOPSの機能が欠損すると、根粒菌はカーリングした根毛の先端から侵入できず、感染糸は形成されない。一方菌根菌は、菌糸を根に侵入させ内生菌糸を伸ばすことまではできるが、そこから樹枝状体を展開できない。感染過程を形態で比べると、根粒菌による根毛への侵入と菌根菌による内生菌糸の伸長とが共通であり、物質のやりとりの上では、共生の最終段階である「バクテロイド」への分化と「樹枝状体」の形成とが共通に見える【図2と図4の比較】。ところが、CYCLOPSの影響は根毛への侵入と樹枝状体の形成に現れたので、話は面倒になった。両者の感染メカニズムの共通現象を正確に対応させるには、CYCLOPSが発現をコントロールしている遺伝子群を同定して、具体的な反応を捉える必要があるだろう。そして、CYCLOPSとCAS/POLという、はからずも根粒共生と菌根共生が関係していることを教えてくれた因子を進化の視点から考察すると、次の一歩を踏み出せるはずである。
5.遺伝子から見た共生の進化
「根粒共生」と「菌根共生」に共通する遺伝子ネットワーク(コモンパスウェイ)は、ミヤコグサのみならず陸上植物に広く保存されている。例外的に菌根共生をしないアブラナ科の代表シロイヌナズナでこれらの遺伝子が見つからないのは、アブラナ科が成立する過程で失われたと考えてよいだろう。一連の遺伝子セットが2つの共生で使い回されていることから、宿主特異的な根粒共生はより一般的な菌根共生から進化したと考えられる。根粒共生特異的な遺伝子(NFRなど)は、菌根共生遺伝子の働きに協調するように派生したに違いない(註2)。根粒共生では、マメ科植物と根粒菌の系統樹により洗練された1種対1種の対応関係(トポロジーマッチ)が存在する可能性もあり、これら根粒共生特異的な遺伝子が深く関わっていると考えられる(註3)。
植物が形成する根粒の中で、根粒菌は細胞膜に包まれたバクテロイドに分化している。この形態はシンビオソーム(共生小器官)とよばれ、プラスチドやミトコンドリアと似ている。プラスチドとミトコンドリアは数十億年前、古細菌にシアノバクテリアや真性バクテリアが共生したものだという説は有名だ。これらのゲノムは、現在では大部分が細胞の核に移行してしまっている。夢のような話だが、根粒菌も遠い将来にはゲノムのかなりの部分を植物の核に吸い取られ、オルガネラになってしまうのではないかと考えることがある。一方、菌根菌は進化の過程で根粒菌と由来を共有はしていても、あくまで一般化の道をとり、ホストとゆるい関係を保ちながら、植物社会の構成に貢献しているようだ(註4)。普段は目にはとまらない、地下で営まれている微生物と植物の共生は、生態系、進化、そして種間の関係などについて様々な姿を見せてくれ、研究に対して多くのアイデアを与えてくれる。
(註3)
一般的に、宿主特異的な関係を築いている生物間では、ホスト・ゲストの系統樹には対応関係(トポロジーマッチ)がみられる。従来はマメ科植物と根粒菌の系統との間にトポロジーマッチはないといわれていたが、最近の研究で、共生に必要な根粒菌の遺伝子を指標に系統樹を書くとトポロジーマッチを示すことがわかった。この理由の1つとして、それら根粒菌の遺伝子群はまとまって菌種間を移動し(水平伝播あるいは水平伝搬)、本来のゲノムとは独立した選択圧と進化を受けてきたことが挙げられる。このようなトポロジーマッチは植物と病原菌のような寄生的な関係において普遍的に見られる現象であり、果たしてマメ科植物と根粒菌の関係は本当に相利共生なのだろうかという疑問も生まれてきた。根粒菌の中にはほとんど窒素固定をせず、植物に依存してぬくぬく生きているものもある。窒素固定はあくまでも「おまけ」で、根粒共生の本来の目的は、植物の光合成産物を奪う片利共生にあるのかもしれない。
(註4)
註2で述べたフランキア共生はそのちょうど中間で、感染1つを例にとっても根粒共生に見られる精巧な認識メカニズムに発展させる前の実験段階であるように見える。フランキアは菌根菌ほどジェネラリストではないが、かといってスペシャリストの根粒菌ほど1:1の宿主特異性を示すわけでもない。フランキア共生に見られるいくつかの感染様式から、優れた系が選択された結果、根粒共生が生まれたのかもしれないとも考えられる。
<謝辞>
共同研究者として多大の協力をいただいた川口正代司、今泉温子、佐藤修正、河内宏、原田久也、Martin Parniskeの各博士にこの場を借りて厚く御礼申し上げる。また、日本のミヤコグサ研究推進に多くの寄与があったミヤコグサコンソーシアムのメンバーにも感謝の気持ちを表したい。この文章を書くにあたっての進化的な考察では青木誠志郎博士に多くを教えていただいた。最後に、ぼくのわがままに付き合ってくれた研究室のメンバーたちのおかげで楽しく研究ができたことを申し添える。
林誠(はやし・まこと)
1966年生まれ。1996年東京大学大学院理学系研究科生物科学専攻修了。博士(理学)。理化学研究所基礎科学特別研究員を経て現在大阪大学大学院工学研究科応用生物工学専攻助手。
.jpg)

.jpg)
.jpg)










