


物理に宿る
生命の紡ぐ物理学
1.生き生きしたカオス
僕の専門は理論生物物理学です。大学入学は1974年で、自然の論理を明らかにする物理が好きで物理学科に進学しましたが、物理によって生きている状態を理解できないかなあと漠然と考えていました。例えばシュレディンガーの『生命とは何か』には、非平衡状態であることの重要性が述べられていましたが、この問題にどのように取り組んだらよいのか具体的な方策はありませんでした。生物物理の最初の授業で、江橋節郎先生が「生物物理には理論なんてありません、実験だけです」とおっしゃって、実験が苦手な僕は大層悲しい思いをしました。ただ、その当時、ノーベル化学賞を受賞したプリゴジンが、非平衡状態の散逸構造で生命を理解しようという学問の流れを主導していて、僕もその影響を受けて、非平衡熱力学と統計力学の研究室へ進み、そこから生物に挑もうと思っていました。しかし始めていくと、非平衡状態の理解が生命の本質につながるのだろうかという疑問も抱いてはきました。

そんな中で、予測不能な系を扱うカオス理論に出会いました。カオスという数学モデルは、初期状態のごくわずかな差が時間の中で大きく増幅されるもので、繰り返しでない運動が生じます。生物のモデルではないのですが、生き生きとしたダイナミックな変化に惹かれました。カオスはミクロな小さな差が増幅しマクロに広がるので、こうした系を集めたらどうなるかということに興味をもち、博士課程からしばらくの間、単純なカオスの要素が、他の要素全部と互いに相互作用させたモデルのシミュレーションを行っていました。これは各要素の集団で決まる全体が各要素の性質に影響するという複雑系のプロトタイプのモデルとなり、多くの新しい現象や概念が見出されました。例えば、全く同じ要素が全体と同じように相互作用しているのに、異なる振る舞いの要素へ分化します。100個の系であれば、そのうち50個が同じ振る舞い、残りの30個がまた別な同じ振る舞い、あと20個はまた別の…といったグループ分けを見せもします。これは細胞分化みたいだなと思いました。多細胞生物では個体を構成している細胞は、みな同じ遺伝子をもち、同じルールで働き得るのに、多様な細胞へと分化するからです。そのあたりから「生きている状態」のダイナミクスを追う、理論生物学の研究を本格化させました。
2. 普遍生物学という物語を
普遍生物学という言葉は、小松左京の50年前のSF小説『継ぐのは誰か?』に出てくる架空の学問の名称です。地球上で進化した生物に限らず、宇宙であろうと、どこであろうと、生物である以上そこに共通するはずの性質を探る学問という設定で、先見の明があったと言えます。物語の中で、普遍生物学を研究する大学院生がコンピュータを使って、生命の可能な状態を調べていて、生物の状態が空間の中でいくつかの領域に引き込まれていくということを発見しています。これはまさに細胞の状態を、力学系の安定した引きこみ先、アトラクターで表現するのとそっくりです。個々の分子によらない普遍性といえば、物理の中で熱力学がもっとも普遍性の高い学問で、これは液体でも気体でもどんな要素であれ安定した平衡状態であれば成り立ちます。そこで「生きている状態」を、そこそこ普遍な系の一つと捉えて、この性質を明らかにすることができれば「普遍生物学」となります。「生きている状態」は、宇宙のどこかの星でも見つかるのではないかと僕は思っていますが、少なくとも地球上ではよく見られる現象です。それは、「非常に沢山の多様な成分が集まって」おり、「まあまあ安定にそれを維持」して、「常にではなくても増える可能性がある」という性質をもった系です。ここで、細胞が倍の大きさに成長して分裂していく際にその組成を維持するには、まず全成分が同じ割合で増える必要があります。この定常成長状態は、平衡状態とも似て、摂動や撹乱によって変化してもほぼ元通りに回復し、相当の範囲まで原状を維持できる頑健性があります。
古澤力さん(理研、東大理物理)との共同研究ですが、何千、何万の成分を持つ細胞を単純化したモデルの計算機シミュレーションを行ってみました。するとこの細胞が安定して成長できるように進化したあとでは、外部環境を変えた時、細胞内部で生じる各成分の変化のそれぞれが独立に増減するのではないことがわかってきました。ある環境で変わりやすい成分は、別の環境でも変わりやすいのです。様々な環境によって各成分の濃度がどう変わったかを空間に表現するとある範囲に拘束されているのです。例えば、成分が5千ある時、その状態は5千次元の空間内を動けるはずですが、実際にシミュレーションして見ると、変化しうる範囲はだいたい一次元の線上、せいぜい二次元の面といった低次元領域に限られていることが見えてきました。膨大な動きうる可能性の中から実際に変われる状態は非常に制限されています。そして、この適応した状態は、撹乱されてももとに戻る傾向をもっています。ただし、どんな変化に対してもただ安定に戻るだけでは外界へ適応した進化が生じなくなってしまいます。このモデルで各成分量の進化的変化を追跡してみると、進化で変化してきた方向には擾乱に対しても変化しやすいということがわかってきました。この方向が先に言った、変化しうる範囲なのです。ここで単に何千成分かがランダムに化学反応して増えているだけの場合ではこうした拘束は生じません。細胞モデルが擾乱に対して安定して増えていくよう進化してはじめてあらわれた性質なのです。
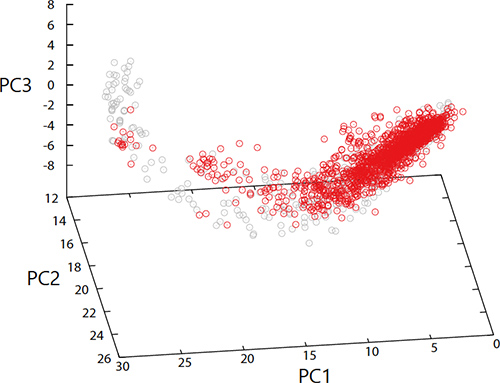
Furusawa & Kaneko 2018 Phys. Rev. E 97, 042410; Kaneko & Furusawa 2018 Annu. Rev. Biophys. 47:273–90)
3. 変わりにくさを支える変わりやすさ
「生きている状態」は、変わりやすさと変わりにくさをうまく共存させているようです。上に述べたように環境条件が変わった時、進化で変化しやすい方向には環境応答やノイズに対してでも変わりやすく、他の方向へは変わりにくいことが見えてきました。古澤力さんの行っている大腸菌を用いた進化実験でも進化で変わる表現型の進化しやすい方向が見えてきています。そして、大腸菌を同一条件下で何回か進化させてもその方向は変わりません。変異自体はそれぞれで異なる場所に入っているのに、です。ナイーヴには生物進化の考えでは、ランダムな遺伝子変化がまずあり、よい表現型が選ばれるとしますが、むしろ表現型は、遺伝的変異よりも環境変化の下で変化しやすい方向が前もってあると考えられるのです。「進化は偶然か必然か」という昔からの問いに、「遺伝子の進化は偶然でも、表現型には前もってある揺らぎやすい方向に拘束される」と答えられるのです。

こうした低次元方向への制約はおそらく発生でも見られます。発生学の浅島誠さんの研究ですが、カエルの個体発生で、初期胚のアニマルキャップと呼ばれる領域から分泌されるアクチビンとレチノイン酸の濃度に応じて、その誘導下に心臓や膵臓などの器官が形成されることが知られています。つまりたった2つの成分の濃度だけで多くの器官が形成されるのです。浅島さんと共同で行った研究で、各器官形成のための分泌因子の濃度の高低と空間配置の順序関係とが対応していることが見えています。さらに発生進化のシミュレーションを行ってみると、やはり表現型変化は束縛されていて、少数の方向にだけが変化しやすく、この方向でのゆっくりとした変化が他の成分の変化を制御しています。この場合もおそらく細胞レベルの現象と、組織、器官を形成する個体発生レベルの現象とが互いにバランスして安定していることが制約となっているのでしょう。
「生きている状態」は、分子ノイズのような微細な時空間スケールの現象から、ゲノムの変化に応じた多細胞生物の進化という長大な時空間スケールの現象まで、ミクロからマクロまでの階層ごとの現象が相互に連関しています。単細胞生物では、分子の複製と細胞の成長との間で、多細胞生物では細胞の増殖分化と組織や器官形成による個体発生との間で、ある種のつり合っている状態を形成しています。こうした階層間の整合性を指導原理として、適応、発生、進化の法則が見え始めていて、その定式化を普遍生物学で進めています。
4. 今、ネオ・ワディントニズム
我々は生物を語る時に、外に現れる表現型、たとえばタンパク質の量は変わりやすいが遺伝子、DNAの配列は変わりにくいと考えますが、こうすると変わりやすいものがまず変化してそのあとで変わりにくいものがゆっくりと変わっていくのが自然です。生物ではこのように速く変化するものと変わりにくいものを共存させているようです。
もし、多くの成分が同じような時間スケールで変化すると、このシステムを、ある特定の方向に変化、進化させることはかなり難しいと思われます。船頭多くして船山に登る、というようにうまく全体が望む方向に変化するのは難しいのです。一方でその中に、少数のゆっくり変わる(変化しにくい)成分があると、その成分が他を制御するようになります。するとその成分を変えれば全体がある方向に動くので、進化しやすくなります。つまり制御側と多数の機能要素に役割分化した系は適応や進化がしやすいのです。一方で次に述べますが進化を通して、こうした非対称的な役割分化も生まれてくるので、役割分化と進化は正の循環構造をもって進みます。

今は進化の捉え方が変わる大きな変革期とも感じています。先にも述べましたが環境の変化に対してまず表現型が変化しやすい方向があり、その後、表現型を固定する役割を遺伝子が果たすことが見えてきています。遺伝子は時間をかけてランダムな変異の中から、適応した表現型を固定化できるものを選んでいく。これは半世紀以上前にコンラッド・ワディントンが提唱した「遺伝的同化」という概念で、今その理論的表現と定量的実証が始まっているとも言えます。20世紀にネオ・ダーウィニズムとして遺伝的進化の総合説が確立したことに倣って言えば、今、我々はネオ・ワディントニズムという形で表現型進化の総合説をつくる時代になっているのかもしれません。それが完成すれば、現存の生きものの変わりやすい性質と変わりにくい性質を測定することで、その生きものの進化の方向と制約性を定量的に予測できるようになるでしょう。
5. 気づいたらセントラルドグマ
最近、ニュージーランドのオークランド大の竹内信人さんと行った研究ですが、まず原始的な細胞を考えて、その中に、自分を鋳型にして複製する分子を設定します。この時、複製のためにはそれを触媒する分子も必要です。今の生物では鋳型情報を担う分子はDNA、触媒はタンパク質と分かれていますが、初期生命ではそのような役割分化は起きていなかったとも考えられます。そこで互いに触媒しあって、複製する分子があったとしましょう。ここで分子の立場としては、自分が他を触媒している間は増殖できないので、触媒機能を失ったほうが増殖には有利です。しかし全部が失ってしまったら誰も増えられません。分子の立場では触媒機能を失ったほうが有利だけれど、細胞の立場で考えれば、そんな利己的分子ばかりでは増えられなくなるわけです。言い換えると分子レベルで有利な方向と細胞レベルで有利な方向が逆行しています。そこでこの触媒と鋳型複製の性質を同じだけもつ2種類の分子からなる細胞の進化シミュレーションを行ってみました、すると、一方の分子種は徐々に触媒機能を失い複製鋳型として働き、他方は触媒機能を失わず鋳型情報の能力を失います。つまり役割分業が、物理でいう対称性の自発的破れとして生じます。この結果、分子レベルの複製と細胞レベルの増殖という、元来逆方向を向いていた要請に折り合いをつけているのです。この役割分化はまさに、DNAは情報を担うけれど触媒機能がなく、タンパク質は触媒として働くけれど鋳型情報の能力がないことに対応しており、分子生物学のセントラルドグマが出現したことになります。多細胞生物での生殖細胞と体細胞、社会性昆虫での女王と働きアリ(蜂)などでも、同様な役割分化が見出されていますが、これも階層間のつり合いから生まれてきたとも考えられます。

機械であれば外部からの設計に従って下位の部品が働けばいいけれど、生物は前もって設計されているわけではありません。部分が集まって上位階層を形成していく際に、階層間で食い違いが生じてしまうかもしれません。それが生じないように、生きている状態を形成していく中で、セントラルドグマや、発生や進化の一般法則が生まれてきたのではないでしょうか。この解明が普遍生物学の課題です。





