

Experiment
神経細胞のパートナー選び
— 標的認識の仕組みを探る。
私たちの体ができるとき,神経細胞から伸びる軸索は,長い道のりを旅して,標的である筋肉や他の神経細胞にたどり着きます。
複雑な体の中で,軸索はどのようにして正しい標的を見つけるのでしょうか。
脳神経系は,神経細胞がもつ軸索と呼ばれる長い突起が他の神経細胞や筋肉などとつながってできている。我々の体の中には,くまなく神経配線のネットワークがはりめぐらされていて,秩序だった行動を可能にしている。たとえば,右手を動かしたいと思ったとき,ちゃんと動かすことができるのは,脳からの指令がいくつかの神経細胞をリレーして,最終的に右手の筋肉を刺激するからだ。
脳神経系のネットワークは,いったい,どのようにしてできあがるのか。思考や記憶,行動の制御など,複雑な機能を司る脳神経系の働きを理解するためには,それができる仕組みを理解することが必要である。
個体の発生過程で神経細胞が軸索を伸ばすとき,たとえ標的細胞が遠く離れていても,神経の軸索は間違いなく決まった道筋にそって伸び,多数の細胞の中から標的を正確に見つけだす。このような神経細胞の驚くべき能力を説明するのに,古くから提唱されてきたのが「標識路仮説」と呼ばれるものだ。
我々が目的地に向かって車を走らせる際,道路標識を確かめながら進んでいくように,神経の軸索もその経路や標的領域に存在する標識を頼りに,その結合の相手を見つけだすというものである。しかしそのような標識が実際存在するのか,存在するならどんな物質の働きによるのか,といったことは不明であった。私たちは,ショウジョウバエの神経系をモデル材料として,この問題に取り組んできた。
ショウジョウバエの神経細胞は,全部で10万個くらいあると予想されている。一見複雑に見えるが,人間に比べれば十分に単純である。なによりも良い点は,ショウジョウバエでは,多くの神経細胞をひとつひとつ識別し,軸索の伸びる様子を追跡できるということだ。たとえば,腹部の筋肉を動かすためのRP1,RP5という隣りあう2つの運動神経細胞は,特定の末梢神経の繊維に沿って軸索を伸ばし,複数の筋肉に向かってそろって進んで行く。ところが標的となる筋肉に近づくと,RP1は必ず12番の筋肉と結合するのに対し,RP5はそれよりひとつ背側の13番の筋肉と結合する(図2)。つまり,これらの運動神経細胞は,何らかの仕組みで自分が結合すべき筋肉を見分けているのである。
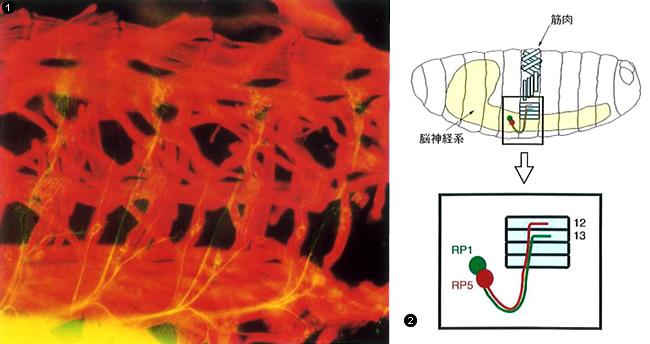
神経細胞を見る
①筋肉に向かって伸びる神経の軸索。4体節分(片側)の筋肉(赤く染まっている)とそこに伸びる神経の軸索(黄色に染まっている)が見えている。
②ショウジョウバエの胚の中で,神経系の組織と筋肉がどこにあるかを示した模式図。一体節分のみがやや詳しく描かれているが,他の体節にも同じように筋肉がある。(神経細胞はRP1とRP5のみが描かれているが,①でわかるようにそれ以外にも多数の神経細胞がある。)
筋肉の上に何らかの標識があるに違いない。しかし,標識の実体についてはわからないことが多かった。そこで私たちは,標識そのものを直接探すのではなく,遺伝子解析の技術を用い,それをつくる遺伝子を探すという方法をとることにした。標識となる分子(タンパク質)をつくる遺伝子は,特定の筋肉において発現することが予想される。そのような遺伝子を,当時開発されたばかりの新しい方法(未知の遺伝子の発現パターンを調べる方法で,エンハンサー・トラップ法という)を用いて探すことにした。多くの遺伝子の発現を調べ,最終的に2つの遺伝子(コネクチンとカプリーシャスと名付けた)を見つけることに成功した。
これらは,もともとは筋肉での発現をもとに見つけた遺伝子だったが,調べてみると,まさにその筋肉と結びつく運動神経においても発現していることがわかった。つまり,それぞれの遺伝子は,異なるセットの運動神経と筋肉のペアーで発現していたのである。さらに,遺伝子がつくるタンパク質を調べたところ,神経や筋肉の細胞の表面にあって,両者の細胞の相互作用に関わる働きをもつことがわかった。つまり,標識として必要な特徴を備えていた。
そこで,より直接的にこれらの遺伝子とそれがつくるタンパク質の働きを調べるために次のような実験を行なった(写真③~⑤)。
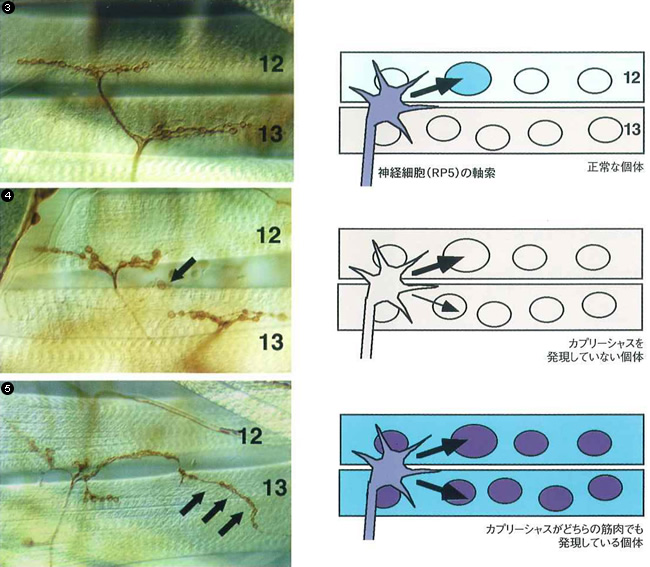
標的を認識するためのタンパク質の働き
正常なハエではRP5という神経細胞は筋肉の12番(カプリーシャス遺伝子からつくられるタンパク質を発現している)に軸索を伸ばす(③)。カプリーシャス遺伝子を働かなくさせたり(④),筋肉の12番と13番の両方で働かせる(⑤)と,PR5からの軸索は筋肉の13番にも伸びてしまう(矢印)。④で,13よりも12のほうに軸索がたくさん伸びているのは,カプリーシャス以外にも,RP5の軸索を12番の筋肉に引き付ける標的があるためと考えられている。
(写真①~⑤および図=能瀬聡直)
カプリーシャス遺伝子は,RP5神経細胞とその標的である筋肉12においては発現するが,隣の筋肉13においては発現しない。そこで,カプリーシャス遺伝子を壊してそれがつくるタンパク質ができないようにしてみた。すると,筋肉12と13の区別がつかないためか,多くのRP5神経は軸索を枝分かれさせて,本来結合しないはずの筋肉13へも結びつくようになった。逆に,本来発現しない筋肉13において人工的にカプリーシャスタンパク質を発現するようにした場合にも,RP5神経は筋肉12と13の両方に結び付いたのである。
以上のことから,カプリーシャスタンパク質が,特定の神経と筋肉がつながる過程に関与していることがわかった。同様の実験によってコネクチン遺伝子のつくるタンパク質についても神経 – 筋結合における標識分子として働くことを示すことができている。
私たちがこの研究を始めた8年ほど前には,神経配線の分子メカニズムはほとんどわかっていなかったが,この数年で爆発的に研究が進んでいる。カプリーシャスやコネクチン以外にも,神経が筋肉を目指して伸び始める初期のころに働くタンパク質など,標識と考えられる分子(タンパク質)が数多く見つかってきている。神経ネットワークの形成の基本的なメカニズムが明らかになり,行動や精神活動の枠組みがどのようにして設計されているのかが,分子のレベルで語れるようになる日もそう遠くないかもしれない。
(のせ・あきなお/東京大学理学系研究科助教授)
※所属などはすべて季刊「生命誌」掲載当時の情報です。
.jpg)
.jpg)











