

Special Story
「翅」が語る生命誌
チョウやショウジョウバエの翅の研究は、私たちの体づくりと無関係ではない。
昆虫の翅とヒトの手足のでき方は基本的なメカニズムが同じであることが明らかになってきた。このことは進化がどのようにして起こってきたのかについても教えてくれる。
美しいチョウの翅を入り口にして、現代生物学が明らかにしようとしていることを見てください。
CHAPTER
最近の研究からわかってきたこと
動物の発生過程は、大きく二段階に分かれる。一つは、前後・背腹の体軸が決まり、それに沿った位置情報に基づいて各器官の元となる原基を生み出す過程。次は、できた原基が独自のプログラムに従って各自独立に形態をつくり出す過程だ。最初の過程では、その器官をつくるのかつくらないのかというもっとも根本的な問題が決定される。つくるべき器官は何かということをこの時期の遺伝子の働きが決めるのである。次の過程では、器官の形態が決まる。したがって、この過程を調べていくと、翅と脚の形がどうして違っているのか、同じ翅でもチョウとショウジョウバエではなぜ形が違うのかといったことがわかってくる。
キイロショウジョウバエでの研究から、体の形を決める基本的な遺伝子の数は意外と少なく、しかもその大部分が、遺伝子発現を制御したり、細胞間相互作用をシグナル分子によって制御したりする制御遺伝子であることがわかった。1万2000あるとされるショウジョウバエの遺伝子のうち、ごく少数に着目すれば形づくりの基本はわかる可能性が出てきたのだ。しかも、それらは制御遺伝子なので、その働きがわずかに変化しても、形に大きな変化をもたらすような進化を引き起こす可能性を秘めている。また、最近ショウジョウバエで発見された遺伝子と、構造的にも機能的にも非常によく似た遺伝子(相同遺伝子)がヒトを含む脊椎動物にも存在することがわかった。そのおかげで、これまでカエル、昆虫など動物種ごとに調べられてきた発生学が遺伝子という共通の言語によって理解されることにより、種の違いを生む進化の問題に取り組めることになった。

ショウジョウバエの翅の成長。卵・幼虫・蛹の赤い部分が翅原基。
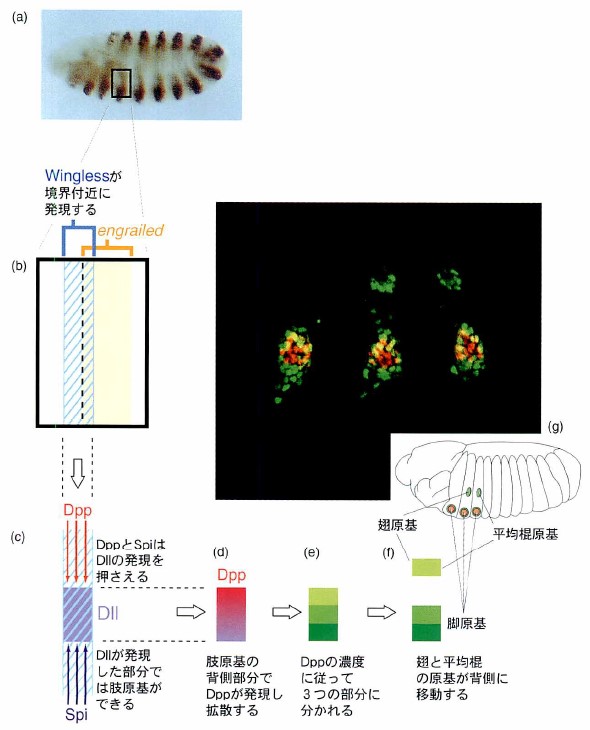
翅原基ができるまで。胚の成長にともない、翅や脚、平均棍の原基ができてくる様子を模式化したもの。受精後5~8時間後の胚(a)。胚はいくつかの体節からできており、体節はさらにengrailedを発現する後部と、発現しない前部に分かれている。肢原基(翅・脚の共通の前駆細胞)の誘導はengrailedの発現部分の前側の境界で起こる(b)。この境界部分で発現するシグナル分子WinglessがDllを発現させ肢原基を誘導しようとする一方で、胚の背側、腹側ではシグナル分子Dpp、SpiがそれぞれDllの誘導を阻害する結果、原基は胚の側方部に誘導される(c)。肢原基の背側にDppが再び発現する。分泌されるDppは濃度の勾配を生ずると考えられる(d)。その濃度の違いをDll発現細胞は感知して、翅、脚近位部、脚遠位部の3種類に分化すると考えられる(e)。翅と平均棍の原基が背側に移動する(f)。
(写真=林茂生)
付属肢の発生メカニズム
では、器官の一つ、付属肢が生み出される過程での制御遺伝子の威力を見てみよう。昆虫の付属肢(翅や脚)は胚の時期(幼虫になるまでの個体)からでき始める。胚には最初、頭や背中の方向を決める軸ができ、この2つの軸に沿った分泌性のシグナル分子の濃度勾配ができる。翅と脚のもとである肢原基はシグナル分子が一定の濃度になる位置につくられる。肢原基は、次に3種類の細胞群に分かれる。そのうち1つは背側方向に移動して翅原基となり、残りの2つは腹側にとどまったまま、同心円状に配列して脚原基となる。ここでもシグナル分子が重要な役割を担っていることがわかっている。このシグナル分子は肢原基の背側に片寄って発現するので、その濃度は腹側にいくほど低くなっていく。肢原基の細胞は、受け取ったシグナル量が多いときには翅原基になり、少ないときには脚原基になるのだ。
こうしてできた原基が翅や脚を形づくっていく(詳しくは多羽田哲也氏のページを参照)。その際重要なのは、前後軸、背腹軸、近遠軸の3つに沿った非対称性がどのようにしてできるのかということである。たとえば翅の前縁部と後縁部は違った形をしているし、背側と腹側も異なっている。また基部と先端部の形も違っている。最近、付属肢の前後や、表裏の違いを決める遺伝子の存在が明らかになった。これらの遺伝子は単に形の違いを決めるだけでなく、性質の違う2つの領域(コンパートメント)を区分し、それらの間でシグナル分子をやりとりさせることで付属肢の成長を制御しているのである。これらの遺伝子はまずショウジョウバエで発見されたが、相同なものがヒトを含む脊椎動物にも存在し、しかも非常に似た空間パターンで発現していることがわかってきた。昆虫と脊椎動物に共通 のメカニズムの存在が明らかになったのである。
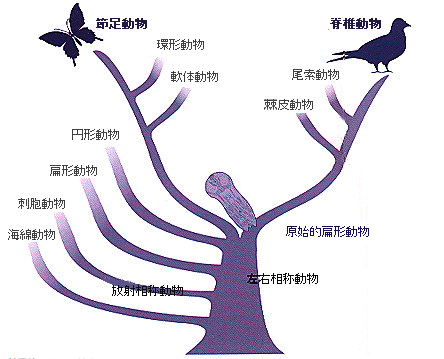
付属肢のルーツは?
動物の系統樹の一例。脊椎動物と節足動物は付属肢をもつ数少ない門であるが、それらの共通 の祖先であると考えられる原始的扁形動物に付属肢はない。
付属肢の進化
では、付属肢は一体どう進化してきたのだろうか。動物は20あまりの「門」に分類されているが、3つの非対称性(前後、背腹、根元と先)がある付属肢を持った生き物は昆虫などの節足動物と脊椎動物にほぼ限られている(イソギンチャクなどの触手は前後、背腹の非対称性を持たないので付属肢には含めない)。ところが奇妙なことに、他の研究結果からは、節足動物と脊椎動物の共通の祖先(左右相称な扁形動物に近いと推定されている)が、ちゃんとした付属肢をもっていたとは考えにくいのである。これは、節足動物と脊椎動物が分岐した後に、独立に付属肢を発達させたことを意味している。おそらく、共通の祖先には、付属肢の発生のカギとなる遺伝子がすでにあったが、節足動物や脊椎動物が出現するまで、付属肢を形成するようには使われなかったのではなかろうか。
現存する動物の「門」は約5億5000万年前のカンブリア紀の比較的短期のあいだに起こった急激な種分化の結果として生まれたとされている。そのときにはすでに発生の基本的な制御遺伝子は存在しており、カギとなるものは、現代のショウジョウバエと同じく少数のものに限られていただろう。カンブリア紀に起こった壮大なドラマは、これらの制御遺伝子が様々に組み合わされることで起こったのではないだろうか?付属肢もまた、このような組み合わせ実験の一つの結果として生まれてきたのかもしれない。節足動物と脊椎動物が類似の付属肢をもつようになったのは、果たして組み合わせの偶然の一致なのか、それとも限られた数の制御遺伝子しか存在していなかったための必然の結果なのかは、今となっては知る由もない。
しかし付属肢の発生の始まり(原基を生み出す最初の過程)をより詳しく探求すれば、付属肢をもたない単純な初期胚の形態から、より複雑な体の体制がつくられる過程は理解できる。そしてこの発生の過程にこそ、付属肢の進化を理解する手がかりが潜んでいるように思われる。進化と比べればとてつもない短い時間スケールで進む個体発生の研究から、ゲノムに刻み込まれた進化の歴史の一端を学べる時代が来ているのである。
(はやし・しげお/国立遺伝学研究所 系統生物研究センター助教授)
※所属などはすべて季刊「生命誌」掲載当時の情報です。.jpg)

.jpg)
.jpg)










