

Special Story
バクテリアから生きものの基本を探る
バクテリアといえども、もちろん種や株によってゲノムはさまざまです。でも、必ずもっているところがあります。複製開始点はその一つ。どんな細胞でもDNAが複製しなければ増殖できないのですから、変わっては困ります。ところで、そこが生きものの面白さ、この複製開始点でさえ少しずつ変化しているのです。さまざまなバクテリアで比べた結果、使う部品は同じで、その数や並び方が違う、いわゆる「使い回し」をしていることがわかりました。「使い回し」は、ゲノムの得意技です。部品の名前などがちょっと複雑ですが、図を見て比べてくだされば、同じところと違うところがわかって面白いと思います。
自然に存在するバクテリアの種の数だけゲノムが存在する。最近のゲノム研究から、すべてのバクテリアゲノムに共通な遺伝子が1000個ほどあるのだが、しかし、それらのゲノム上の配置はさまざまだということがわかった。進化の過程で自由に動きまわり、変化したらしい。ところが変化しない場所が1ヵ所存在する。それが、私たちが研究を続けてきた複製開始点領域だ。
1963年、バクテリアの環状ゲノムの複製は決まった場所から両方向に起こることがわかり、特定のDNA領域(レプリケータ)に特定のタンパク質(イニシエータ)がつくと複製が始まるという仮説が提唱された。それから十数年後、大腸菌ゲノムからレプリケータとイニシエータの遺伝子が単離され、レプリケータは245塩基対で、その中にイニシエータが結合する9塩基対の配列(TTATCCACA)が5個存在すること、イニシエータはDNA二重鎖の巻き戻しを誘発するタンパク質(DnaA)であることがわかった。
その頃、私たちのグループは進化的に大腸菌とは遠縁の土壌バクテリア・枯草菌ゲノムの複製開始点を決めようとしていたのだが、なかなかうまくいかなかった。その理由は後でわかる。その時は、ともかく複製開始点を含むと考えられる広範囲の塩基配列を決めるという攻め方で、なんとか開始点を推定した。今のように配列決定が自動化される前のことだったので、約1万塩基対の決定にはかなり時間がかかり、配列データベースが存在しない状態での情報解析は至難の業だった。しかし、幸い得たものは大きかったので、苦労のし甲斐はあったと思っている。
じつは心の中では、まったく違う種なのだから、様子がだいぶ違うだろうと予想していたのだが、それに反し、イニシエータはDnaA、その結合配列も大腸菌とまったく同じで、さらにdnaA遺伝子周囲の十数個の遺伝子配置まで共通とわかった。その後の研究で、枯草菌だけでなく、すべてのバクテリアに共通とわかってきた。
さて、もう少し詳しく複製開始点領域を比べてみよう。レプリケータが開始点として働くには、DnaAタンパク質の結合配列(DnaA-box)が存在するだけでは不十分だ。複数のDnaA-boxが、特定の位置と向きに配置される必要がある。このような領域全体を複製起点(origin of Chromosome)という意味で、oriCと呼ぶ。大腸菌のoriCは前述したように245塩基対の中にDnaA-boxが5個配置された単純な構造をしていた。ところが枯草菌のDnaA-boxは15個もあり、しかも3領域(各々145、626、191塩基対)に分散し、領域間にはイニシエータdnaA遺伝子も挿入されるという、なんとも複雑な構造だった。その後にわかったシュードモナス菌のoriCはdnaA遺伝子を挟んで2領域に、ストレプトミセス菌は枯草菌と同じ3領域に分散していた。私たちが枯草菌から複製開始点を大腸菌のように簡単に決められなかったのは、予想外の長さ(全長2400塩基対)と複雑な構造のためだったのだ。
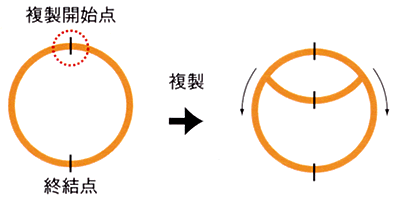
バクテリアの環状ゲノムの複製は決まった場所から両方向に起こる。
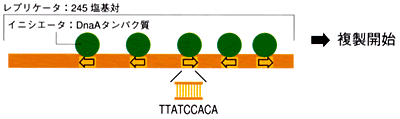
大腸菌K-12のレプリケータ(=特定のDNA領域)にイニシエータ(=DnaAタンパク質)が付くと複製が始まる。
わかってみると大腸菌は変わり者で、大部分は複数のDnaA-box領域をもち、その配置はdnaA遺伝子を挟んだ2~3領域に分断されていたのである。このようにゲノムの複製開始点を担い制御する二つの要因のうち、イニシエータタンパク質(DnaA)は共通だが、oriCは共通のDnaA-boxを核としながらもゲノム上で多様に変化している。したがって、DnaAタンパク質が結合して生じるDNAの構造変化もさまざまということなのだろう。
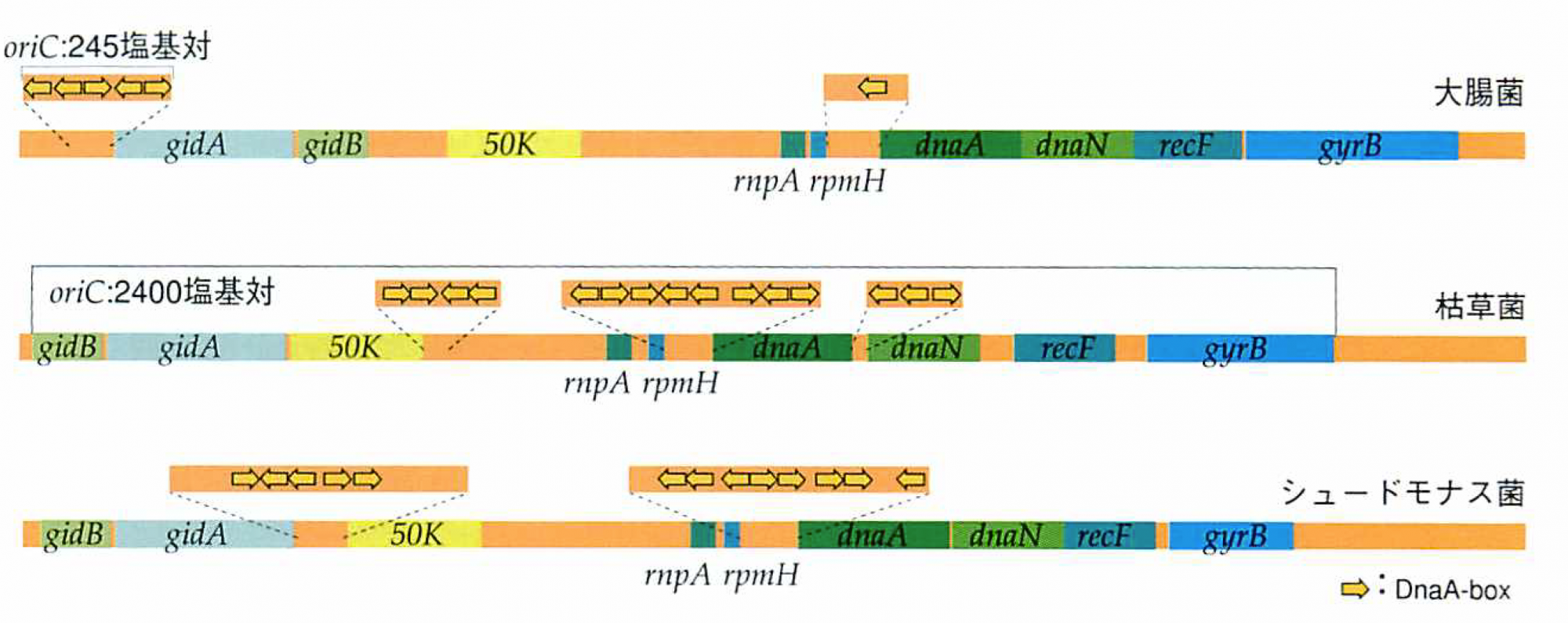
3種のバクテリアの複製開始領域
大腸菌のoriCは245塩基対の中にDnaA-boxが5個配置された単純な構造。枯草菌のDnaA-boxは15個で3領域に分散し、領域間にはイニシエータdnaA遺伝子が挿入され、全他は2400塩基対に及ぶ。シュードモナス菌のDnaA-boxはdnaA遺伝子をはさんで2領域に分散している。dnaA遺伝子周辺領域を調べると、10数個の遺伝子配置がバクテリアにほぼ共通であることがわかった。
バクテリアの複製開始点領域は大腸菌型、シュードモナス型、枯草菌型の3種類に大別できる。それらから共通項を求めると、原始細胞のoriCはdnaA遺伝子と少数のDnaA-boxとが隣接していたものだと想像できる。十数億年にわたる進化の過程で、dnaA遺伝子はほとんど変化することなく機能を保存し、DnaA-boxの塩基配列にいたってはまったく変化していない。変わったのはDnaA-boxを含む領域の数と配置、領域の中のDnaA-boxの数である。枯草菌を含むグループであるグラム陽性菌に進化した仲間では、多くのものが領域を3倍化し、そのうちの2個をoriCとして使っている。一方、グラム陰性菌では領域を倍化し、そのどちらか、あるいは両方をoriCとして使っている。そのなかで大腸菌など寄生バクテリアとして進化した腸内バクテリア群では、2領域の一部のDNAが逆位を起こし、oriCをdnaA遺伝子から5万塩基対ほど遠ざけ、残った領域からDnaA-boxを除いてoriC機能をなくしたのだ。すなわち、oriCのようなシグナルとなるDNAも、遺伝子とまったく同じに重複、欠失、転位によって多様化していることがわかる。
このようなoriCの構造の多様化は、開始点の働きを多様化しているのだろうか。大腸菌のようなdnaA遺伝子から離れた単純なoriCは自律性が強く、同じ細胞の中で複数共存できる。寄生バクテリアのように栄養豊富なところでどんどん増殖するにはoriCがたくさんあってもよいという性質は有利なのだろう。一方、枯草菌など複雑なoriCはdnaA遺伝子の発現と強く連携しており、開始タンパク質の要求度が高く、同種のoriCを排除する負の調節機構になっている。これは、栄養飢餓状態(自然界ではよくある)で栄養細胞から胞子細胞へ切り替えて待ちの姿勢に入る時の制御にも使われているらしい。
複製開始点領域はゲノムの中でもっとも保守的でほとんど変化していないように見える。しかし詳細に解析すると、共通の素子を上手に使って、さまざまに構造を進化させ、バクテリアの多様な生き方にみごとに対応していることが見えてきた。この「使い回し」はこれまでにもゲノムの特徴とされてきたが、「バクテリアのDNA複製開始」という生命現象のもっとも基本の場でこれが明確になったことは興味深い。
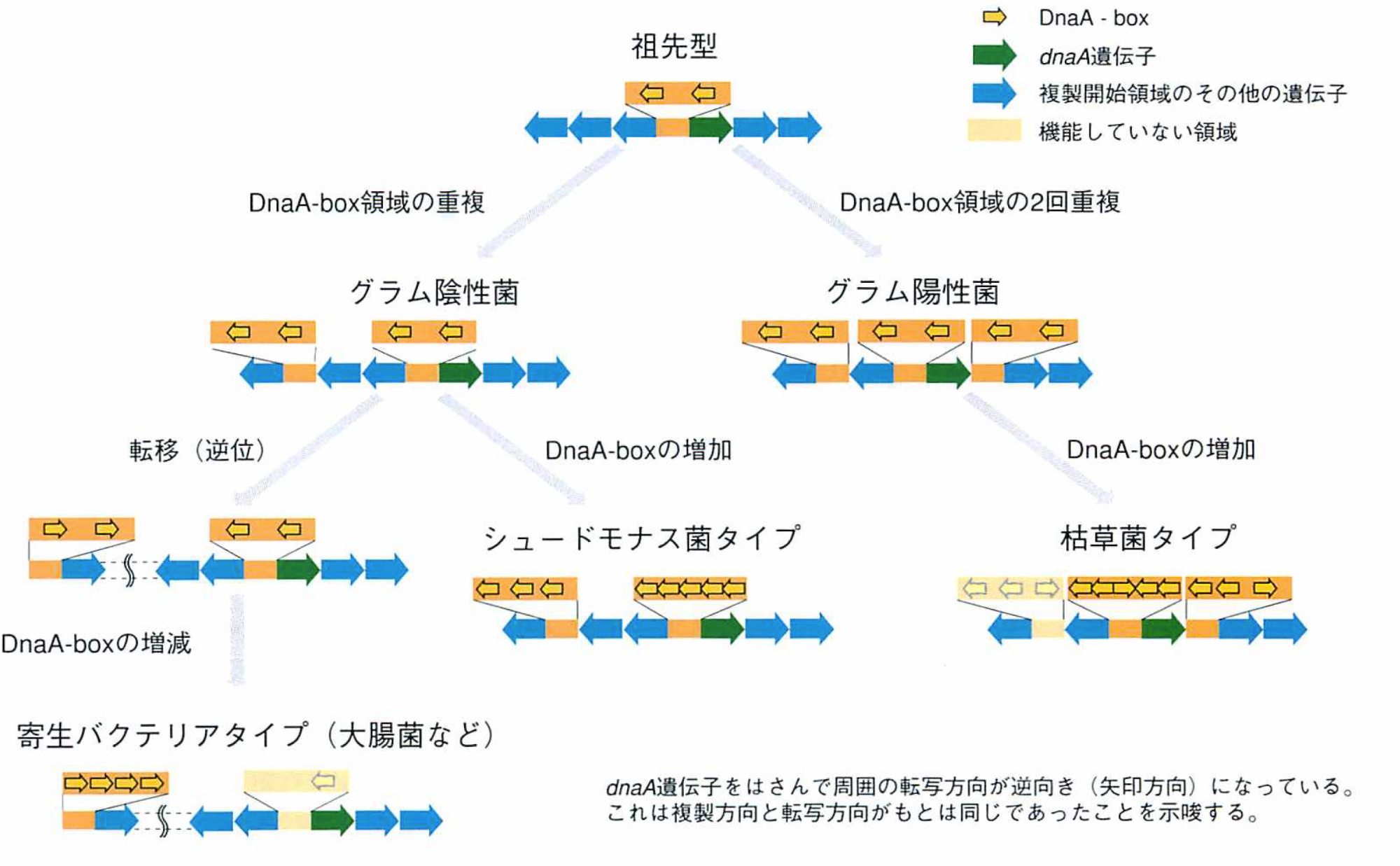
oriCの進化モデル
DnaA-boxを含む領域の数と配置、領域の中のDnaA-boxの数が変化し、3タイプのoriCが生じたと考えられる。原始のoriCはdnaA遺伝子と少数のDnaA-boxとが隣接していたのだろう。枯草菌を含むグラム陽性菌は、領域を3倍化し、そのうちの2個をoriCとして使うようになり、グラム陰性菌では領域を倍化し、そのどちらか、あるいは両方をoriCとして使っている。その中で大腸菌など寄生バクテリアとして進化した腸内バクテリア群では、2領域の一部のDNAが逆位を起こし、oriCをdnaA遺伝子から5万塩基対ほど遠ざけ、残った領域からDnaA-boxを除いてoriC機能をなくしている。
吉川 寛(よしかわ・ひろし)
1933年神戸市生まれ。JT生命誌研究館顧問。東京大学化学系大学院生物化学専攻終了、理学博士。カリフォルニア大学バークレー校助教授および準教授、金沢大学がん研究所教授、大阪大学医学部教授、奈良先端大学院大学教授、およびバイオサイエンス研究科長を歴任、2001年より当館へ。研究分野はゲノムの複製とその制御、真核生物の細胞周期、微生物ゲノムの構造と機能。蝶を愛するモレキュラーナチュラリスト。
.jpg)

.jpg)
.jpg)










