

RESEARCH
進化と生態のつながりを示す周期変動
<食う-食われる>の関係にある生きものは、互いに影響をあたえることで、周期的に変動すると考えられます。小さな生態系の観察で、これまでのモデルでは説明できない振動パターンを発見しました。迅速な進化と生態系現象のつながりを探ります。
1.個体数はどう変わる?
「自然の世界に生きる生物たちの個体数は、自然のバランスによってほぼ一定に保たれているのではないか」という直観は、実は当たっていないことが多い。個体数が突然増えたかと思うと、しばらく後に急激に減り、しかもゼロになることはなく(絶滅することなく)存続するという例がよく見られる。
個体数変化のパターンは、大まかに「平衡」「周期変動」「カオス」の3つに分けられる。「平衡」は一定の個体数が続く安定した状態、「周期変動」はある周期で増えたり減ったりする繰り返しパターン、「カオス」はでたらめに変動しているように見える複雑な変化パターンである。野外での長期にわたる観測例は多くはないが、平衡やそれに近い状態が7割、周期変動が3割の生物で見られ、カオスははっきりとは認められないことがわかってきた。
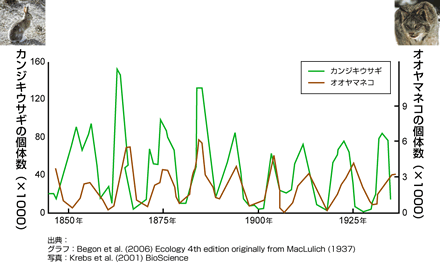
(図1) カンジキウサギとオオヤマネコの周期変動
カナダの森林にすむカンジキウサギとその捕食者であるオオヤマネコは、10年周期の個体数振動を見せる。世界でもっとも詳しく研究された個体数振動の一つである。これまでの研究により、この2種の個体数振動は2種間の捕食ー被食関係が原因ではなく、他の多様な生物を巻き込んだ複雑な関係によることがわかってきている。
ある一定の周期で増えたり減ったりをくり返す周期変動は、まるで機械の仕業のようにも見える(図1)。なぜ、バランスが取れているように見える平衡状態をやぶって、個体数の振動がおきるのだろうか。それを引き起こすしくみの一つが、捕食-被食(食う-食われる)の関係である。被食者が増えると、それを餌とする捕食者が追いかけるように増える。すると被食者は捕食者に食われて減少し、やがて餌不足にみまわれた捕食者が減少し、難を逃れた被食者がふたたび増加する。この周期変動を説明する数学モデルが出されている(図2)。
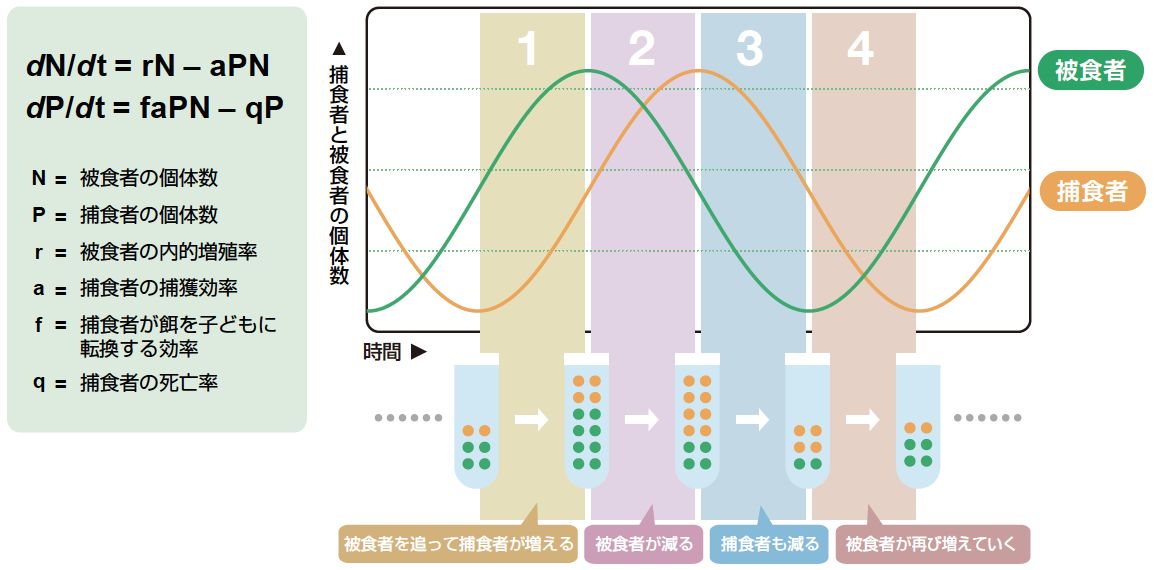
(図2) ロトカ-ボルテラの捕食モデル
捕食者と被食者の周期変動が捕食ー被食の関係により形作られることを再現するもっとも基本的な数学モデル。捕食者と餌生物の個体数変化が微分方程式により表される。この数式にしたがって個体数が周期的に変動し、1周期は4つの状態に分けて理解できる(右図)。
2.迅速な進化が変える相互作用
同じ種でも、他の個体より多くの子孫を残す個体もいれば、なかなか死なない個体もいる。そうした違いが遺伝的にきまる場合、進化を引き起こす原動力となる。進化とは、生物集団の遺伝的組成が時間的に変化することであり、原理的には1世代でも起こりうる。つまり、現在の世代と次の世代の遺伝的組成が異なれば、それは進化と呼ばれるのである。
これまで生物の進化といえば、種分化や適応放散に見られるような、長い時間をかけて起こる現象が注目されてきた。しかし、薬剤耐性細菌のように人間が強い選択圧をかけることで短時間でも進化が起きることは知られているし、実は、自然環境の中でも迅速な進化が起きうることが見つかっている。ダーウィンフィンチはおよそ15世代の時間の中で嘴の大きさや形・体の大きさに迅速な進化が観測された例である(図3)。同じ適応進化といってもさまざまな時間スケールの進化が存在するのだ。
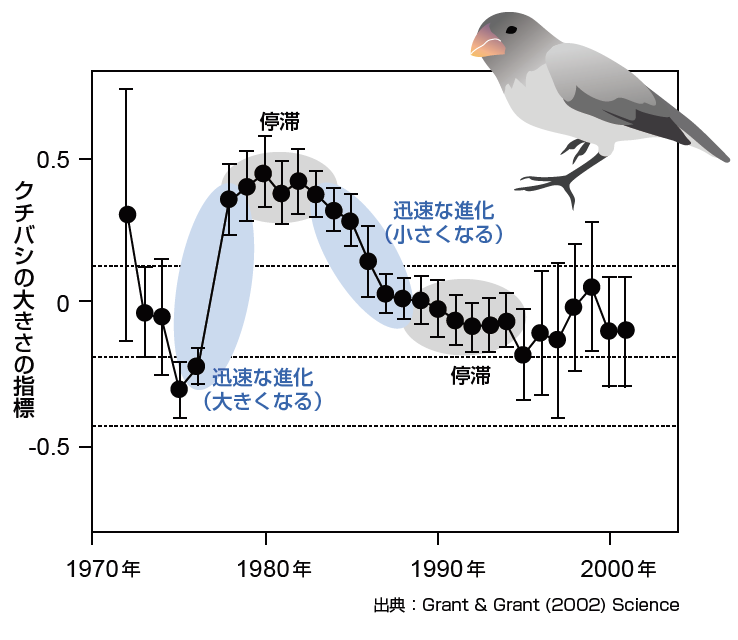
(図3) ダーウィンフィンチに見られる迅速な進化
ガラパゴス諸島での30年にも及ぶ長期観察の結果、フィンチの形質(この図ではクチバシのサイズ)が生息環境の餌条件に応答して迅速な進化を見せることが発見された。迅速な進化は必ずしも一定方向に継続して起こるのではなく、停滞する期間や反対方向への進化も見られる。
迅速な進化による形質の変化は、その性質が関係する個体間の相互作用に変化をもたらし、個体数の変化を引き起こす可能性がある。捕食-被食の関係の場合、防衛形質が迅速に進化した被食者は捕食者に食べられにくくなり(捕食-被食の相互作用が変化し)、捕食者が増えてもそれほど影響をうけないだろう。これまで、個体数の変化パターンと迅速な進化を関連づけて考えることはほとんどされてこなかったが、両者は密接に関係しているはずである。
3.迅速な進化を引き起こす遺伝的トレードオフ
実は、私たちも最初から個体数の変動と迅速な進化を結びつけて考えていたわけではない。当初は自然湖沼のプランクトン群集での個体群動態を知りたいと思ったのだが、そこには複数の捕食者と被食者が存在し複雑な食物網をつくっているため、短期間での変化を説明することは難しい。そこで、淡水プランクトンであるワムシ(Brachionus calyciflorus)と、その餌となる藻類のクロレラ(Chlorella vulgaris)をケモスタット(註1)というミクロコズムの一つで飼育し、詳細な個体数の変化を調べた(図4)。

(図4) 〈ワムシークロレラ〉のケモスタット培養系
内部には新鮮な培養液が常時流入しており、栄養塩を吸収したクロレラが増殖し、それをワムシが捕食して増殖する。流入と流出は同じ速度で行われ、余った栄養塩・クロレラ・ワムシが系外に流出する。
クロレラは無性生殖で増えるため、子孫は同じ遺伝子型をもつクローンと考えられる。そこで、実験に用いたクロレラ個体群は単一のクローンから成ると漠然と考えていたのだが、その場合にあてはまる数学モデルでは説明できない個体数の変化パターンが現れた。なぜだろう。集団内に複数の遺伝子型が存在すると仮定すれば、環境の変化によって遺伝子型の頻度が変化し(迅速な進化が起こり)、その結果、個体群動態が大きく変化する可能性がある。そこで、この仮説を前提として、遺伝子型を調べたところ、実験に用いたクロレラ個体群には捕食者であるワムシに対して防衛能力の異なる形質をもった遺伝子型が複数存在することがわかった。しかも、これらの遺伝子型間(クローン間)で、防衛能力と競争能力の間にトレードオフが存在するのである(図5)。そこで、ワムシが多くクロレラが少ないときには、防衛タイプが適応的になるが、ワムシが少なくクロレラが多いときには、競争能力の強い非防衛タイプが有利になってその個体数を増やすと予測される。つまり、ワムシとの関係で異なる遺伝子型が選択されると考えられるのである。
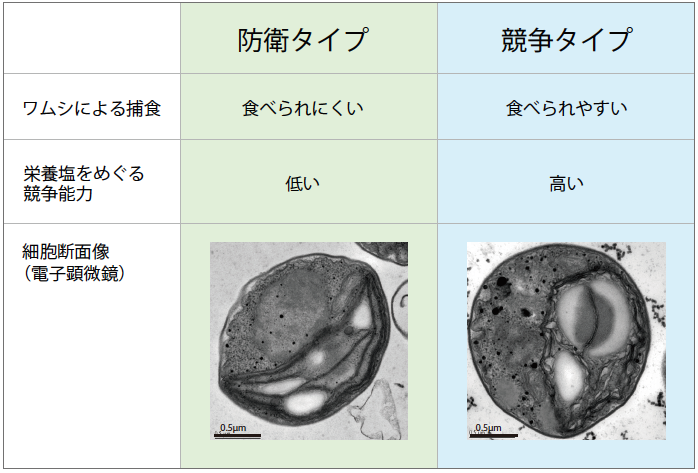
(図5) クロレラの遺伝子型にみられるトレードオフ
ワムシによる捕食されやすさと、栄養塩をめぐる競争能力の間にはトレードオフがあり、どちらか一方の性質でしか、より適応的にならない。細胞断面画像は極めて似ており、クローンの違いをもたらす構造はまだわかっていない。
(註1) ケモスタット
連続的に培養液を交換し、プランクトン生物を飼育する装置。長期間にわたって人工的に制御された環境で飼育が可能で、微分方程式を用いた個体群動態の記述に取り組みやすいことから、個体数変化を研究するのに格好のモデル系として古くから用いられてきた。
4.多様性がもたらす進化と生態のフィードバック
私たちは、個体数の振動が変わったことと、被食者であるクロレラの迅速な進化とを結びつけて理解できるはずであるという仮定から出発している。それを確かめるために、迅速な進化が有るときと無いときでの個体数の変化を比較する実験をした。幸運なことに、クロレラは無性生殖で増えるため、一つのクローンを培養すれば単一の遺伝子型をもつ個体群を、遺伝子型が異なるクローンを合わせて培養すれば、複数の遺伝子型をもつ個体群をつくることができる。前者には遺伝的多様性がないので迅速な進化は起こらない(ただし、突然変異によって新しい遺伝子型が侵入するまでは!)が、後者には遺伝的多様性があり迅速な進化の条件が満たされる。この性質を利用して、迅速な進化が個体数の変化パターンを変えるかどうかを調べた(図6)。
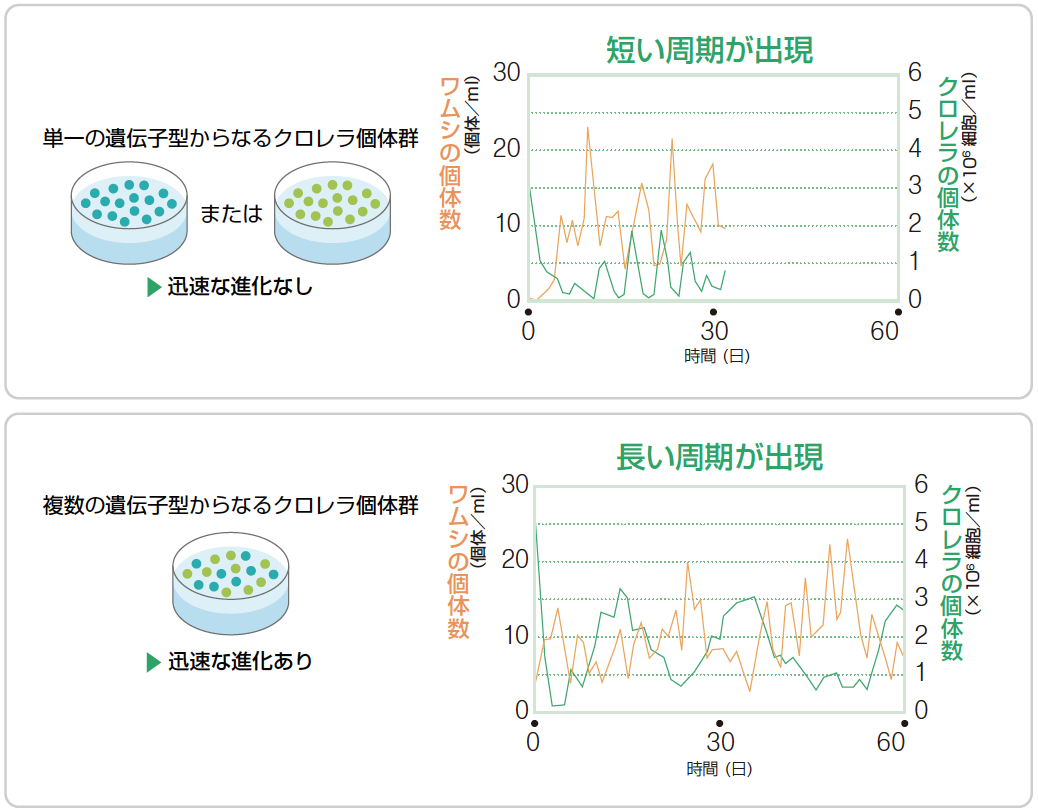
(図6) クロレラの遺伝的多様性と個体数の振動パターン
クロレラ個体群の遺伝子型が単一の場合は短い周期(上)、複数の場合は長い周期(下)が現れた。
同一の遺伝子型をもつクロレラを単離し、ワムシと一緒に培養して個体数の変化を観測すると、約6-10日という短い周期の振動が見られた。また、クロレラの振動とワムシの振動のずれは周期の約25%であり、これはロトカ-ボルテラのモデルが示す典型的な捕食-被食の関係の個体群振動と一致する。一方、複数の遺伝子型からなるクロレラ個体群を用いて、同様に個体数の変化を観測すると、約30-60日という長い周期を見せ、クロレラの振動とワムシの振動のずれは周期の約50%でちょうど逆位相の関係にあった。両者の違いはクロレラ個体群に遺伝的多様性が存在するか否かなので、後者ではクロレラが迅速に進化し、それが個体数振動のパターンに影響したと考えられる。
この結果は、遺伝的多様性が存在することで適応進化が引き起こされ、それが個体数の振動パターンを変化させ、さらには個体数変化が自然選択をもたらす原動力となるというフィードバックがあることを示している。さらに詳しく調べると、ワムシの密度と栄養塩濃度の変化に応じて、異なる性質をもつクロレラの遺伝子型(クローン)の頻度が変わることがわかった。ワムシが多く捕食圧が高いときには防衛タイプが高まり、ワムシが少なく栄養塩を巡る競争が激しいときには、競争タイプの頻度が高まったのである。
5.振動をつくるのは量より質
先の実験と同じようにクロレラに遺伝的多様性がある条件で個体数振動を観測してみると、これまで観測したことがない奇妙な個体数振動も見られた(図7)。生態学の理論では、捕食者の数が増えれば被食者の数は減少すると考えられてきたが、捕食者のワムシの個体数が大きく変動するにも関わらず、餌のクロレラの個体数はほとんど変化せずほぼ一定だったのだ。ケモスタット内でのクロレラとワムシは密接な捕食-被食関係にあるはずなのに、個体数振動のパターンだけを見ると、この2つの生物の間にはあたかも相互作用がないかのように見えるのである。従来の捕食者-被食者系の個体群動態をあつかった理論では説明のつかない現象である。
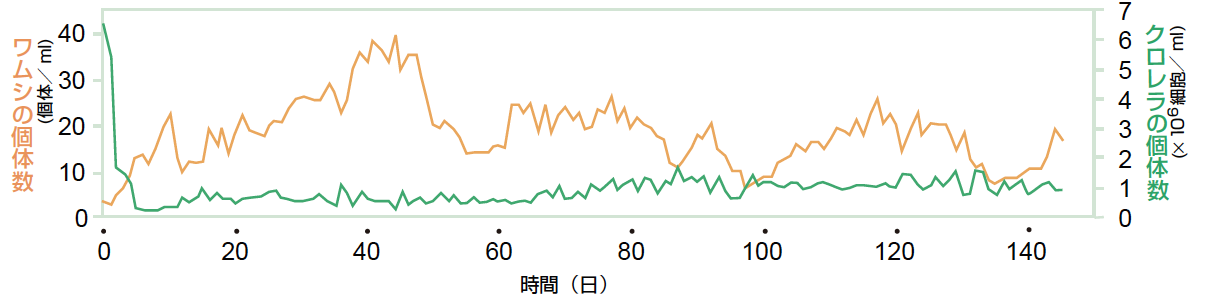
(図7) 奇妙な個体数振動
餌生物の個体数はほぼ一定なのにも関わらず、捕食者の個体数は大きく振動していた。
実験的な検証は今後の課題であるが、数理モデルをもちいた理論研究ではこの奇妙な個体数振動を説明することができた。ワムシの個体数が大きく振動するのに対応して、防衛タイプと競争タイプのクロレラの個体数がちょうど逆位相に変化する振動パターンを考えればよいのである。この場合、クロレラの個体数は互いに補償するために(競争タイプが多いときは防衛タイプは少なく、防衛タイプが多いときは競争タイプは少ない)クロレラ全体の個体数はほとんど変化しないことになる。ワムシの個体数に大きな振動をもたらしたのは、クロレラの個体数ではなく、クロレラの平均的な性質(食べられやすさ)だったのである。この奇妙な個体数振動を「隠された個体数振動」と呼び、実験による検証を試みている。
6.個体数と自然選択の「めぐる」
私たちの研究は、個体数の周期的な振動に見られる「めぐる」と、状況が変われば異なる遺伝子型が選ばれるという自然選択に見られる「めぐる」が、相互に密接に関係していることを明らかにしてきた。自然界では多くの生物個体群が個体数変動を見せており、その中にはたとえば水産生物の漁獲量の変化など、人間生活に関係の深い生物も含まれる。前述のフィンチの例からも、自然での環境要因の移り変わりは、自然選択圧の変化を比較的簡単につくりだしているようである。
実際、一方向への迅速な進化は多くの場合は長続きせず、自然環境では行ったり来たりの「めぐる」自然選択(振動選択)が多いと考えられている。野外生物の個体数変化パターンを理解するにあたって、私たちの研究が明らかにした迅速な進化の影響は無視できないだろう。餌となる藻類の迅速な進化が、小さな生物群集の生態的な現象(個体数の変化パターン)をがらりと変えてしまうのだ。
個体数変化や生物群集の組成のような「生態的な現象」と、さまざまなスケールで起きる「進化的な現象」は、互いに密接に関係しており、その一つの側面が迅速な進化と、その個体数変化への影響であった。「生態的な現象」と「進化的な現象」の相互関係の理解はまだ始まったばかりであり、さまざまな因果関係が近い将来明らかにされるだろう。短い時間の中で起こる進化に注目した私たちの研究は、個体数変化のメカニズムを理解するための礎の一つを提供していると考えている。
吉田丈人(よしだ・たけひと)
2001年京都大学生態学研究センター(理学研究科博士課程)修了。博士(理学)。米国コーネル大学ポスドク研究員・リサーチアソシエイト、日本学術振興会海外特別研究員・特別研究員、東京大学総合文化研究科講師を経て、2008年より同准教授。
.jpg)

.jpg)
.jpg)










