

RESEARCH
液胞へと荷物を運び容れる
植物細胞の中のしくみ
液胞は植物細胞の約9割を占める巨大なオルガネラ(細胞小器官)だ。植物にしかないこの独自のオルガネラはどのようなしくみによって成立しているのだろう。必要な時に、必要な場所へと物質を輸送する細胞内のしくみから液胞の営みを知る。
1.真核細胞の膜交通と多様なオルガネラ
私たち真核生物の細胞内には脂質の膜で区画された多数のオルガネラ(細胞小器官)が存在する。小胞体はタンパク質の合成と品質管理を行い、ゴルジ体はタンパク質の加工と仕分けを行うなど役割の異なるオルガネラが連携して細胞が生きることを支えているのだ。ところで、真核生物はオルガネラをもたない原核生物から派生したとされている。オルガネラの由来は2通りある。ひとつは細菌が細胞内に取り込まれてオルガネラ化する細胞内共生で、ミトコンドリアや葉緑体はこうして生まれた。その他のオルガネラは細胞膜が内側にくびれこみ、区画化したものだと考えられている。この区画が徐々に分化し多様なオルガネラとなったのである(図1)。
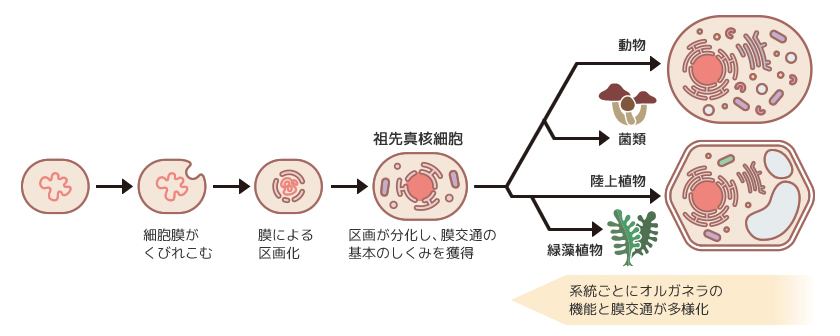
(図1) 真核細胞のオルガネラと膜交通の多様化
オルガネラの機能に必要なタンパク質や脂質など多くの物質の組成はオルガネラごとに異なっている。そこで、細胞内にはオルガネラ間で物質を輸送するしくみがあり、膜でできた小胞や細管を介して物質をやりとりするので「膜交通(メンブレントラフィック)」と呼ばれている。輸送の出発地となるオルガネラではタンパク質など物質の選別と小胞や細管への積み込みが行われる。目的地のオルガネラではまず小胞と目的地の膜の結合(繋留)がおこり、続いて膜同士が融合して積荷が下ろされる。この一連の過程が、様々なオルガネラや細胞内外の境界である細胞膜の間で起こることにより、精緻な膜交通のしくみができあがっているのだ。
真核生物の進化の過程で系統ごとにオルガネラの機能が多様化したので、10億年以上前に分岐した動物と植物とでは役割の異なるオルガネラがいくつか存在する。それぞれの系統で新しいオルガネラが生まれ、古いオルガネラとは別のはたらきを獲得してきた過程はわかっていない。また、オルガネラの機能や性質が変化していった時、そこにどのような分子のはたらきが介在したのかも未解明である。しかし、新しいオルガネラ機能を成立させるには、そこへ物質を運ぶ膜交通経路の改変が必要であったことは間違いなかろう。私は植物の膜交通のしくみの解明を通してオルガネラの多様化の道のりを探っている。
2.植物独自のオルガネラ・液胞
新たなオルガネラのはたらきが生まれてきた過程を探る材料として植物の液胞は非常に優れている。液胞の役割は動物のリソソームと同じく不要物を分解したり、オートファジー(註1)を介して栄養分を再利用することであり、菌類の液胞はまさにその機能をもっている。ところが植物の液胞はそれに加えて独自の機能をもっており、そのはたらきが私たちの生活とも密接に関わっている。そのひとつは様々な物質の貯蔵である。植物性タンパク質の代表格と目される大豆タンパク質の多くや、お米のタンパク質の一部は液胞の中にある。また果物に甘さや酸味をもたらす糖や有機酸、花びらに鮮やかな色彩をもたらす色素も多くが液胞に蓄積されている。植物が屋久杉やメタセコイアのように動物と比較して、大きくなることができるのは液胞が大量の水を貯めることで空間を充填し細胞を大きくする役割をしているからである。このような植物の液胞の機能の成立には、多様な物質を大量に運び込むしくみが必要なはずである。植物の液胞輸送経路は他の生きものの液胞やリソソームへの輸送経路に比べて機能強化されているのではないだろうか。私たちは植物の液胞への輸送のしくみの特徴を探るため、膜交通を支える分子を調べた(図2)。
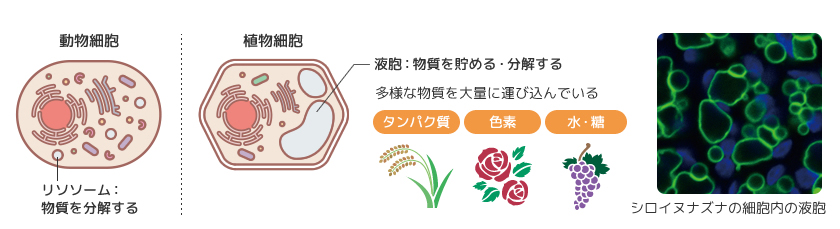
(図2) 植物の大きな液胞の多様な役割
写真(右)は蛍光顕微鏡でとらえたシロイヌナズナの細胞。
緑:液胞膜、青:葉緑体の自家蛍光
(註1) オートファジー
自食作用とも呼ばれる、細胞内の不要タンパク質分解のしくみ。2016年、オートファジーの仕組みの解明により大隅良典博士がノーベル生理学・医学賞を受賞した。
関連記事:
・季刊「生命誌」62号「自分を食べて生き残る細胞に魅せられて」大隅 良典
・季刊「生命誌」45号「オートファジーの現場をとらえる」水島 昇
3.膜交通に関わる分子たち-植物独自の分子の発見
膜交通の基本は全ての真核細胞で共通している(図3)。まず、出発地で荷物の選別と詰め込みが行われ、膜が出芽して小胞となる。目的地に辿り着くまでに複数のタンパク質が連携してはたらいており、これらを制御するのが小胞膜上のRABタンパク質だ。RABが活性化すると、小胞は細胞骨格がつくる道を運ばれ、目的地の膜に繋ぎとめられる。その後、膜と膜を引き寄せるSNAREタンパク質によって膜同士の融合が起きるのである。
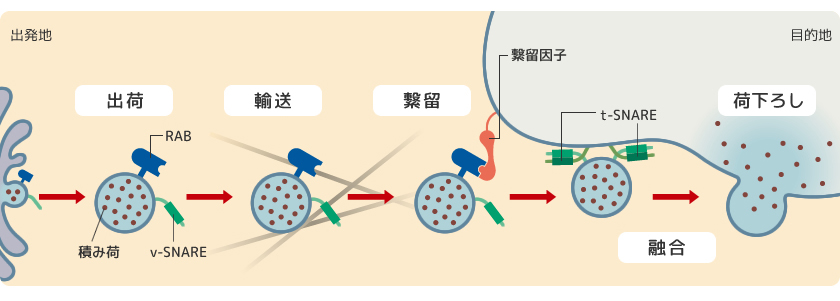
(図3) 膜交通の基本のしくみ
出発地となるオルガネラでは積み荷の選別・濃縮が行われ、輸送小胞の出芽が起こる。この時、輸送小胞の膜にはRABとv-SNAREが結合している。RAB活性化後に細胞骨格がつくる道を運ばれた輸送小胞は繋留因子によって目的地の膜に繋ぎとめられる。その後、輸送小胞のv-SNAREと目的地の膜のt-SNAREが結合し互いに巻きつくことで2つの膜の融合が起こり、荷が下ろされる。
膜交通ではたらく分子の構造は動物と植物の間で非常によく似ているのだが、私たちはシロイヌナズナを用いて植物がいくつか変わり者の分子をもつことを見出した。そのひとつがSNAREの一種、VAMP727だ。このSNAREは、動物細胞がもつVAMP7というSNAREの仲間だが、タンパク質のアミノ酸配列中に酸性の挿入配列をもつ(図4)。被子植物のゲノムには他の生物と比較して非常に多くのVAMP7の仲間がコードされており、VAMP727に加え、2つのグループVAMP71、 VAMP72が存在する。その中で動物のVAMP7と似ているVAMP71と植物独自のVAMP727が両方とも液胞膜ではたらいていることがわかった。液胞膜で膜の融合を実行するSNAREが複数あることが植物の液胞輸送経路の強化に役立ったのではないかと考え、私たちはさらに液胞輸送経路の実態を追った。
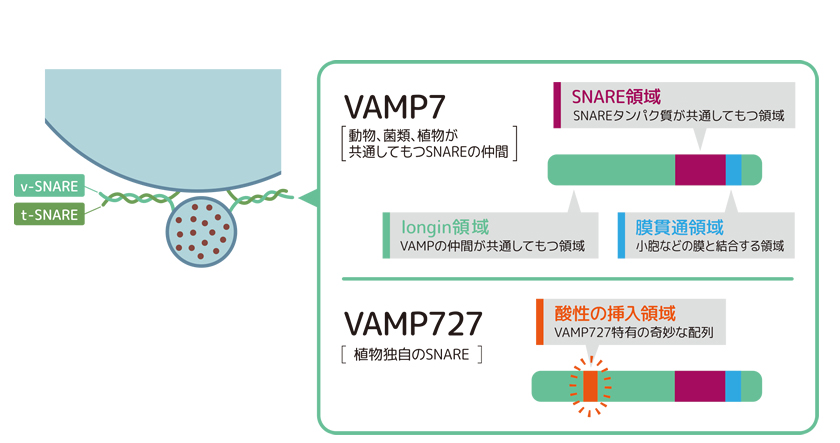
(図4) 植物独自のSNARE・VAMP727
VAMP727はVAMP7によく似ているが、一部だけ特有の配列をもつ。図では分子を模式化して描いているが、実際にはt-SNARE は3種類(Qa、Qb、Qc)または2種類(Qa、 Qb+c)の分子によって構成される複雑な複合体だ。
4.2つの分子スイッチが順番にはたらく巧妙な輸送経路
酵母と動物の細胞ではRABについての解析によって、オルガネラのひとつであるエンドソームを介した液胞・リソソームへの輸送経路が明らかになっている(図5)。2つの分子、RAB5とRAB7が順番にはたらいてエンドソームの性質を変化(成熟)させることにより、液胞・リソソームへと物質を輸送する経路である。まずRAB5と繋留因子CORVETがはたらき初期エンドソームに複数の小胞が繋留されて融合する。大きくなったエンドソームではRAB5からRAB7への付け替えが起こり、こうして成熟した後期エンドソームが液胞・リソソームへと向かう。後期エンドソームでRAB7とともにはたらくのは繋留因子HOPSである。この一連のしくみには複数の宅配物を配送センターでまとめてから目的地に向かうのと同じ巧妙さがある。この経路ではたらく分子をシロイヌナズナで包括的に調べた結果、同じ経路が植物にも存在していることがわかった。これは動物、菌類と植物が共通の祖先に由来することを示すと考えてよかろう。
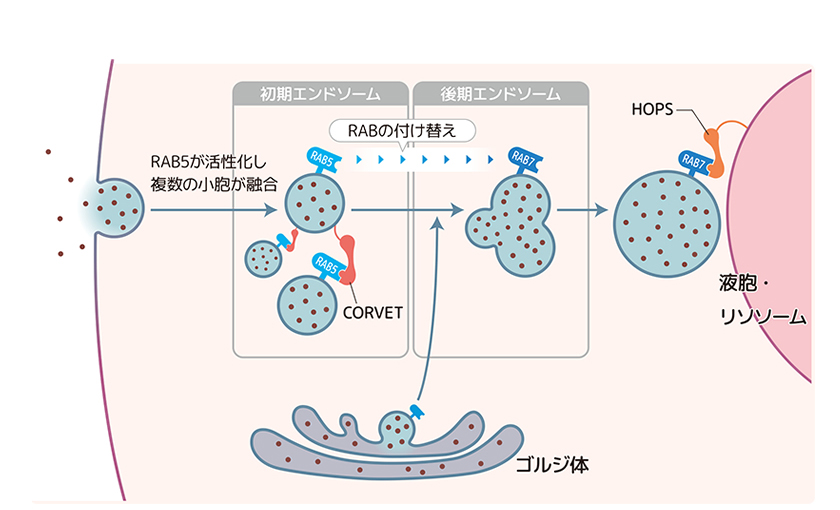
(図5) 菌類や動物の液胞・リソソームへの輸送経路。
5.古いものと新しいものの上手な組み合わせ
ところで植物には、祖先由来の輸送経路に加え、RAB5だけがはたらき、RAB7が不要な液胞輸送経路と、RAB5とRAB7がどちらも不要な液胞輸送経路が存在していることがわかった。RAB5だけがはたらく液胞輸送経路はこれまで他の生きものでは見つかっておらず、植物が進化の過程で独自に獲得したものだといえる。そしてこのRAB5だけが必要な輸送経路で液胞膜を融合している分子こそ、VAMP727だったのである。さらに私たちは、RAB5とVAMP727が関わる輸送経路でCORVETがエンドソームと液胞の繋留を担うことも見いだした(図6上段)。これらの結果は、植物が進化の過程で新たに獲得したVAMP727を、祖先から受け継いだRAB5やCORVETと組み合わせることによって、植物細胞に新しい液胞輸送経路ができたことを示している。こうして多くのタンパク質が液胞へと輸送可能になったのであろう。
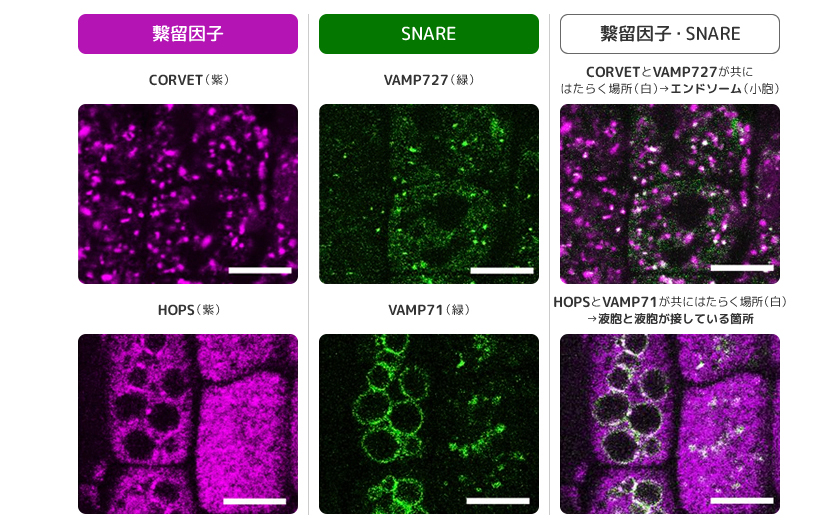
(図6) 膜交通の分子がどこではたらくかを見る実験
細胞内のタンパク質を蛍光で標識し、どこではたらいているかを見た。スケールバーは5 μm。
写真の出典:Takemoto, K et al., Proc Natl Acad Sci USA(2018) 115: E2457-E2466,
https://doi.org/10.1073/pnas.1717839115
液胞と液胞は融合して巨大化することがある。ここではRAB7とHOPS、VAMP71がはたらいていることを私たちは見出した(図6下段)。シロイヌナズナでCORVETやHOPSの機能を阻害したところ、種子がきちんと形成されず育たなかった。大豆やシロイヌナズナは種子形成時に多量のタンパク質を液胞に貯め、これを発生の際の栄養に使うことが知られている。必要な時に、必要な荷物を液胞へ運ぶことは植物が生きていくうえで必須のしくみなのだ。
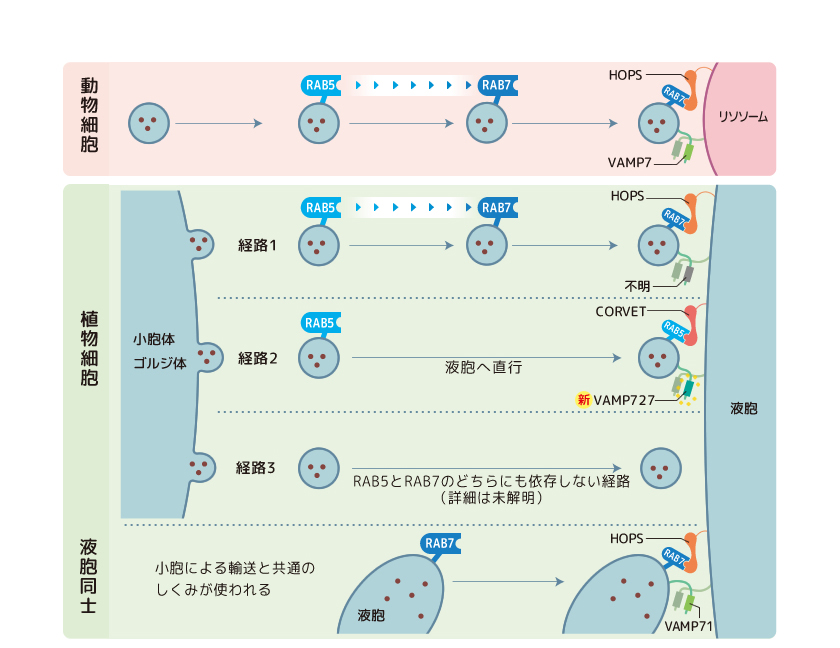
(図7) 見えてきた植物の液胞への輸送経路
植物は動物と共通の輸送経路に加えて、独自の経路を2つ獲得したようだ。独自経路は分子の使い方を変えたり、新しい分子を獲得することで開拓されたと考えられる。
私たちは植物における液胞輸送の多様化の一端を明らかにできたが、まだ多くの問いが残っている。例えばVAMP727はどのようにして他のVAMP7の仲間とは異なる仕事ができるようになったのか。植物は独自の液胞輸送経路をいつ獲得したのか。そもそも、オルガネラの新生はどのようにして起こったのか。これらの問いを明らかにしたいと、私たちはシロイヌナズナの解析に加えて、陸上植物の中で最も初期に分岐したコケ植物の仲間であるゼニゴケの解析も進めている。植物での解析結果と動物、菌類の知見を重ねることで、オルガネラと膜交通の多様化の道のりを明らかにし、そこから植物独自の生存戦略を捉えたいと考えている。

上田 貴志(うえだ・たかし)
1998年東京大学大学院理学系研究科生物科学専攻博士課程修了。博士(理学)。理化学研究所研究員、東京大学大学院准教授、JSTさきがけ研究員(兼任)などを経て2016年より基礎生物学研究所教授。
.jpg)

.jpg)
.jpg)










