

RESEARCH
消えた塩基と増えた色
ネコのオレンジ毛ができるしくみ
ネコの毛の色を黒かオレンジに決定する遺伝子がX染色体上にあることは、100年以上前から知られていた。しかし、長らくその遺伝子の実体は未解明だった。解明の鍵を握っていたのは、「塩基配列の欠失」。DNAの変異によって広がりを見せたネコの毛の色に注目してみましょう。
1. 三毛猫はメスばかり
皆さん、三毛猫はメスばかりであることをご存知でしょうか。これは、3色の毛のうち、黒かオレンジかを決める遺伝子がX染色体上にあることと関わっているためで、サビ猫も同様です。白についてはオス・メスの区別なく決まる別のしくみがあり、のちほど少し触れます。オスはX染色体を1本しかもたないため、黒かオレンジどちらかになりますが、メスはX染色体を2本もっており、黒とオレンジ両方の遺伝子をもつことができます。しかし、メスの各細胞ではどちらか1本が働き、残る1本はその働きが抑制されます(図1)。毛に色素を供給する色素細胞の中で、どちらが選ばれるかは発生過程でランダムに決まるのです。この「X染色体のランダム不活性化説」を最初に唱えたのは遺伝学者のMary F. Lyonで、1961年のことでした。
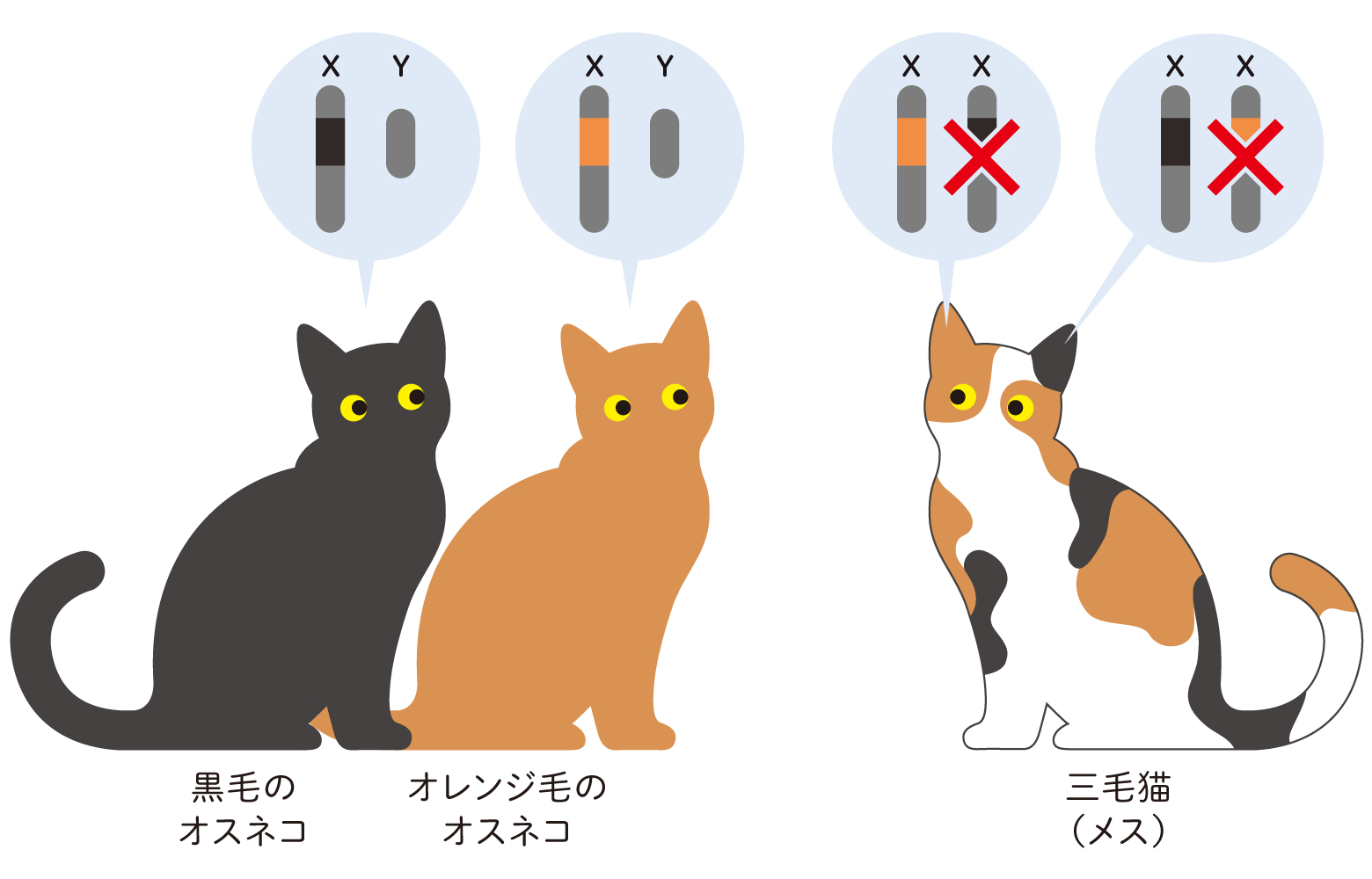
(図1) メスのX染色体不活性化
左:オスはX染色体を1本しかもたないため、黒とオレンジが混ざった模様はできない。
右:メスはX染色体を2本もっており、胚発生の過程で、細胞ごと、領域ごとに、どちらか一方がランダムに不活性化する。
以来、これが定説として広く社会にも浸透することとなりました。しかし、X染色体上にあるとされ、オス・メスを問わず少数派であるオレンジの毛をもたらす遺伝子の実体は、ゲノム解読が盛んに行われている今日に至っても、依然不明のまま。いくら待てどもこの謎解きに挑む研究者は現れませんでした。「では、自分たちで解明しよう!」と私たちの研究が始まりました。
2. 「オレンジ遺伝子」の実体は?
先人たちの研究により「オレンジ遺伝子」がX染色体上のどのあたりにあるかは、大まかに絞り込まれ、その領域に遺伝子が12個あることまでは明らかにされていました。名前がわかっている遺伝子については、すでに情報が蓄積されているヒトやマウスなどのデータを参照することで、その働きをある程度推し量ることができます。ところが12個の候補遺伝子のリストには、毛色や色素合成に関する遺伝子の名前がありませんでした。ネコでは、いずれかの遺伝子が、マウスやヒトでは知られていないやり方で毛色の決定に関わっているのでしょうか?
ネコは実験動物ではありませんから、生体から採取した血液や皮膚の小片などからゲノム配列は解読できても、実験で、個々の遺伝子の働きや表現型への影響を確かめるようなことはできません。
私たちは、まず福岡市内の獣医さんが診療で採取したネコの血液の一部を、飼い主の了解を得た上で譲り受けました。そして、三毛猫やサビ猫など模様の異なる個体でDNA配列にどのような違いがあるのか比べてみることにしました。18匹のネコの全ゲノムの配列を決定し、それらのデータを国立遺伝学研究所の中村保一教授らが作成した高精度イエネコ※ゲノム配列にマッピングしました。18匹のネコの内訳は、三毛が8、サビが1、オレンジ毛が1、それに比較対照としてオレンジ毛をもたないネコが8匹です。
※私たちが日常、親しんでいるネコは生物学的にイエネコ(Felis catus)と分類されます。
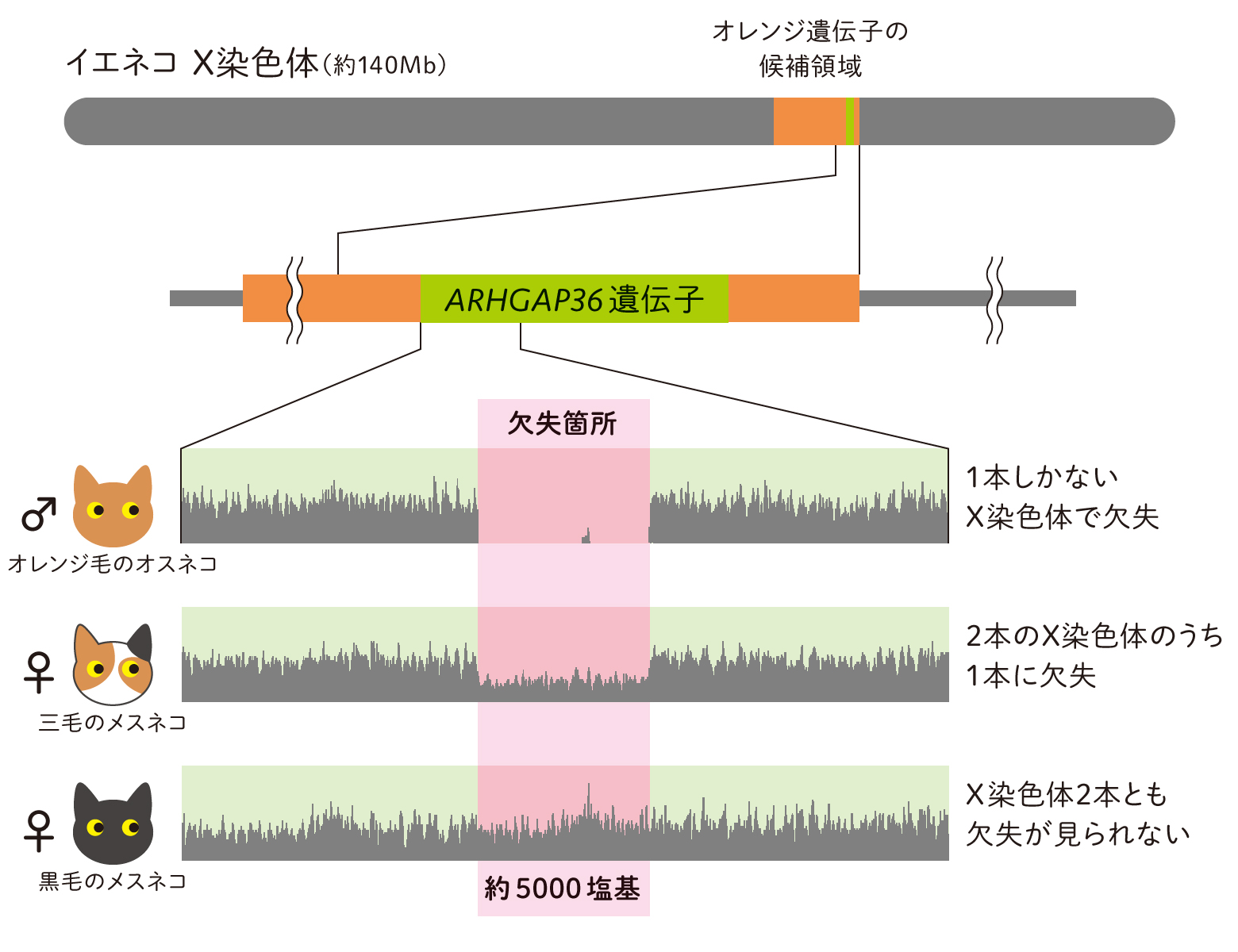
(図2a) オレンジ毛をもつネコに特有の変異(欠失)を発見
解析の結果、12個の候補遺伝子のうちのARHGAP36遺伝子(Rho GTPase-Activating Protein 36)に、オレンジ毛をもつネコにのみ約5,000塩基の欠失が確認されました(図2a)。欠失していたのは、ARHGAP36遺伝子の発現を抑制するように働く塩基配列でした。通常、遺伝子の近傍には、転写を促進する因子が結合するエンハンサーと呼ばれる配列や、反対に、転写を抑制する因子が結合するリプレッサー又はサイレンサーと呼ばれる配列があります。今回、欠失が確認された配列はリプレッサーだと思われました。この配列がなければ細胞内でARHGAP36遺伝子の発現が活発になるはずです。どうやらこの遺伝子が、毛色を黒からオレンジに変える現象に関わっているようです(図2b)。
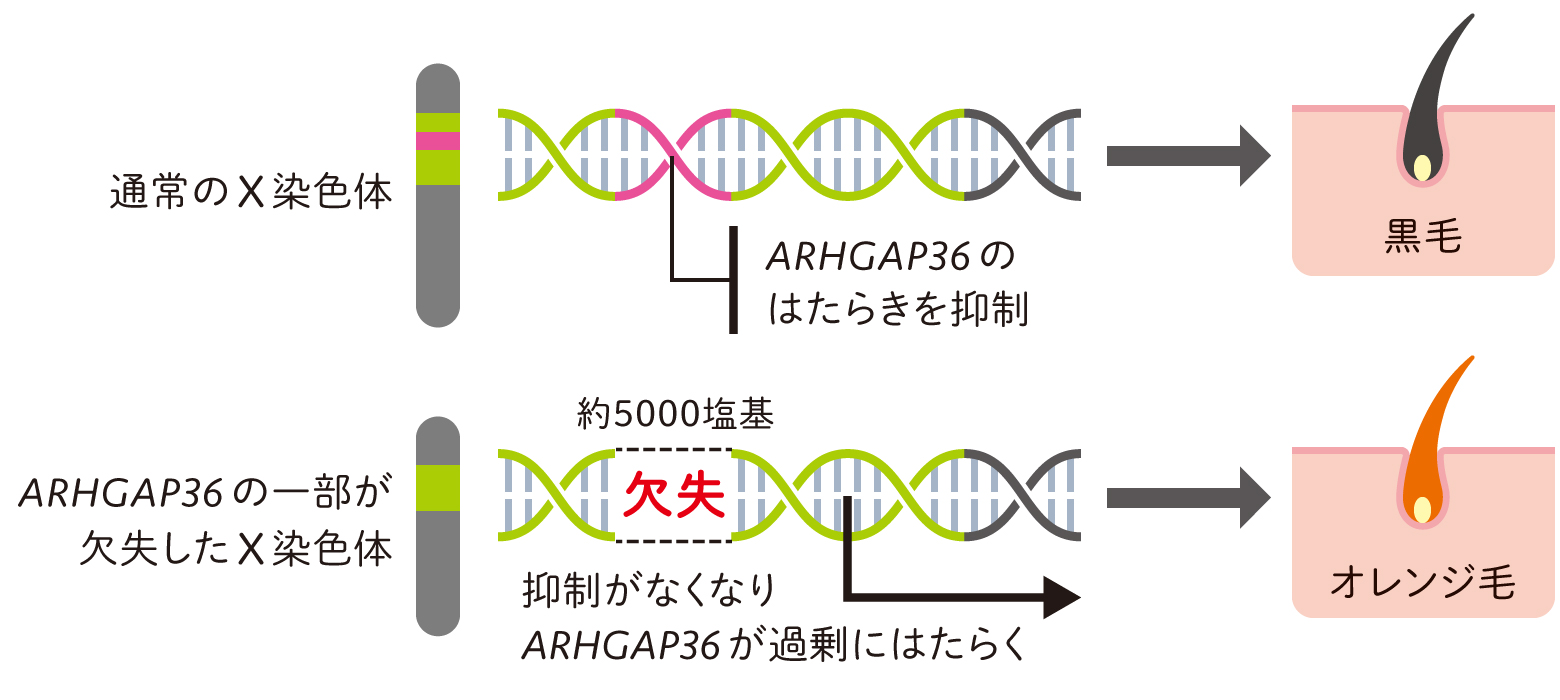
(図2b) 遺伝子を抑制する塩基配列の欠失により毛色が変化する
3. 変異によって毛色が変わるしくみ
では、白い毛はどのようにできるのでしょうか。体毛に黒やオレンジの色素を供給している色素細胞は、私たち脊椎動物で、初期胚の頃、頭部背側で生じる神経堤細胞に由来します。色素細胞は神経堤細胞から分化しつつ、その分布を背側から腹側へ徐々に広げていくので、体中に色素細胞が行き渡れば全身に色がつきます。しかし、三毛猫などがもつ白毛をつくる遺伝子はこの遊走に影響し、色素細胞が到達できない領域が白く残ります。三毛猫でも背側や顔の上部に色がつき、腹側に白い領域が多いのはこうした理由です(右写真)。この白毛の遺伝子は常染色体にあるので、性別とは関係しません。


ARHGAP36遺伝子は、神経堤細胞に起源をもつ一群の細胞系譜で発現しており、細胞内のシグナル伝達を調節するプロテインキナーゼA(PKA) に結合して、その働きを抑制することが知られています。最近、ヒトのARHGAP36遺伝子が、先天性皮膚疾患や皮膚がんの原因遺伝子であることも報告されました。
そこで、細胞内のさまざまな代謝や化合物の反応経路についての情報を網羅したデータべース:KEGG(Kyoto Encyclopedia of Genes and Genomes)を参照して、ヒトやマウスではどのように色素合成が誘導されるのかを確かめました。
色素は、アミノ酸のチロシンを出発点として、酵素チロシナーゼ(TYR)などの反応により色素細胞にあるメラノソームという袋状の小器官の中で合成されます。黒い色素はユーメラニン、オレンジの色素はフェオメラニンという化合物で、前者の合成経路には、TYRのほかチロシナーゼ関連タンパク質1(TYR1)やドーパクロームタウトメラーゼ(DCT)などの酵素も関わっています。これらの遺伝子の上流でシグナル伝達を担うのがPKAです(図3上)。細胞内でARHGAP36遺伝子の発現が上昇すると、ARHGAP36タンパク質が結合してPKAの分解を促進するため、その下流の働きが低下すると考えられます(図3下)。
この経路がフルに働くと主にユーメラニンを合成しますが、オレンジ毛をもつネコの場合、ARHGAP36の発現増加によってPKAが媒介するシグナル伝達が阻害され、フェオメラニンが合成されるようになるのだろうと考えられます。
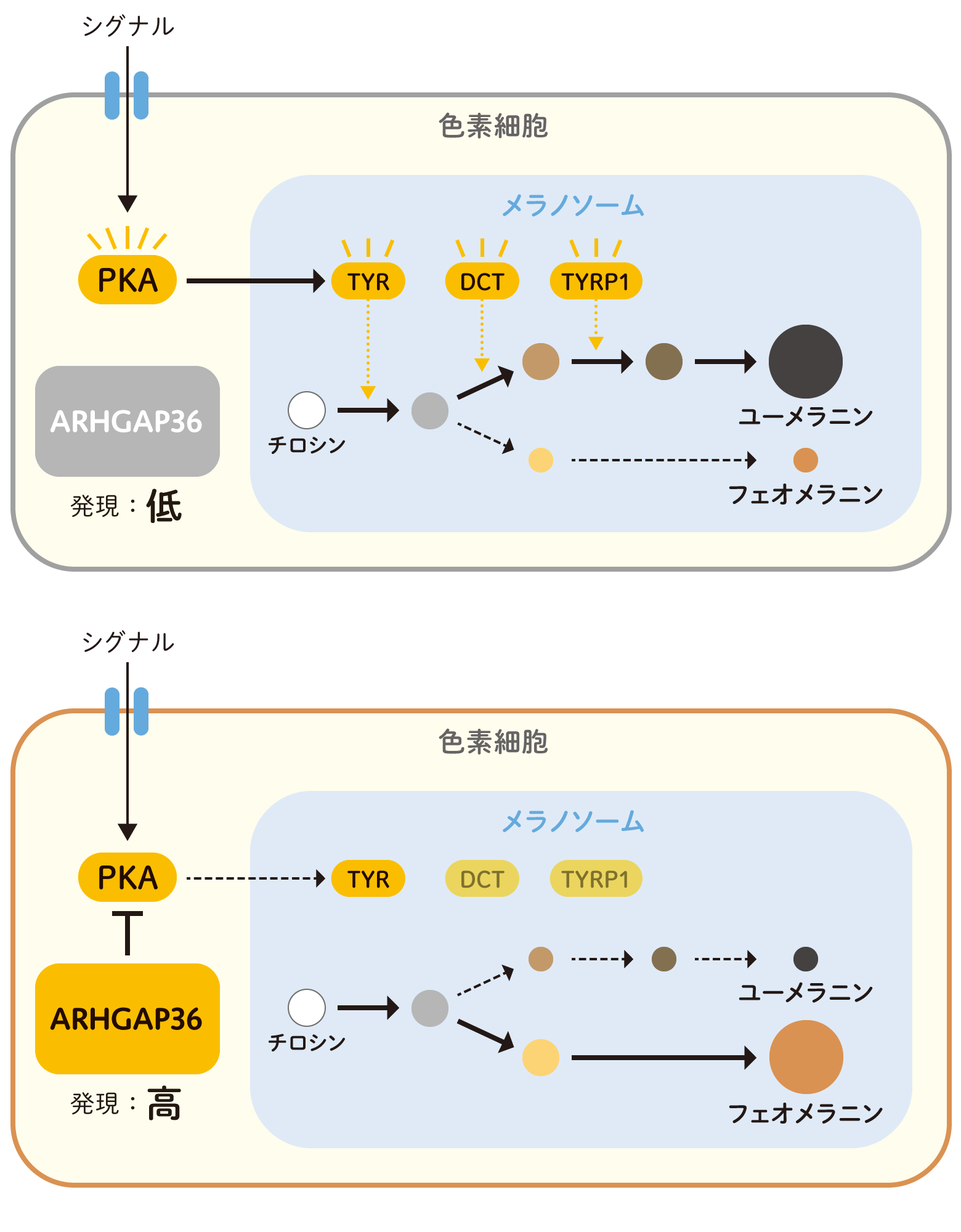
(図3)色素細胞のメラノソーム内で色素がつくられるしくみ
上:PKAの下流で活性化する色素合成に関わる酵素TYR, DCT, TYRP1の反応により、メラノソーム内で、アミノ酸のチロシンから2種類の色素ユーメラニンとフェオメラニンが合成される。この経路は通常、主にユーメラニンを合成している。
下:ARHGAP36の発現によりPKAの活性が抑制されると、ユーメラニン合成に関わる経路の働きが低下し、フェオメラニンをより多く合成するようになる。これは、本来ユーメラニンへ変換されるべきだった中間化合物が蓄積し、フェオメラニン合成に使い回されるためだと考えられる。
4. X染色体不活性化の不思議
ここでもう一度、X染色体不活性化について確認しましょう。この現象においては、遺伝子の抑制にDNAのメチル化が大事な役割を果たします。DNAの塩基A・T・G・Cのうち、C(シトシン)にメチル基が修飾されると、転写因子がDNAに結合できずに遺伝子の発現が抑制されるのです。特に塩基配列でCとG(グアニン)が連続するCpGアイランドと呼ばれる領域でメチル化(Me)が多く起こります(図4)。
まずは公開されているヒトとマウスのデータを再解析し、2つあるX染色体のうちのどちらか一方で、ARHGAP36遺伝子が不活性化されていることを確認しました。次に私たちのネコのサンプルを用いて、DNAメチル化の有無と分布について測定と解析を行いました。そして、オスとメスでARHGAP36遺伝子のCpGアイランドのDNAメチル化状態を比較したところ、メスにのみX染色体不活性化の目印となるメチル化が確認されました。同様のメスに特異的なメチル化は、マウスでも確認することができました。つまり、ネコでもヒトやマウスと同様に、X染色体不活性化が起きていることが推測されました。
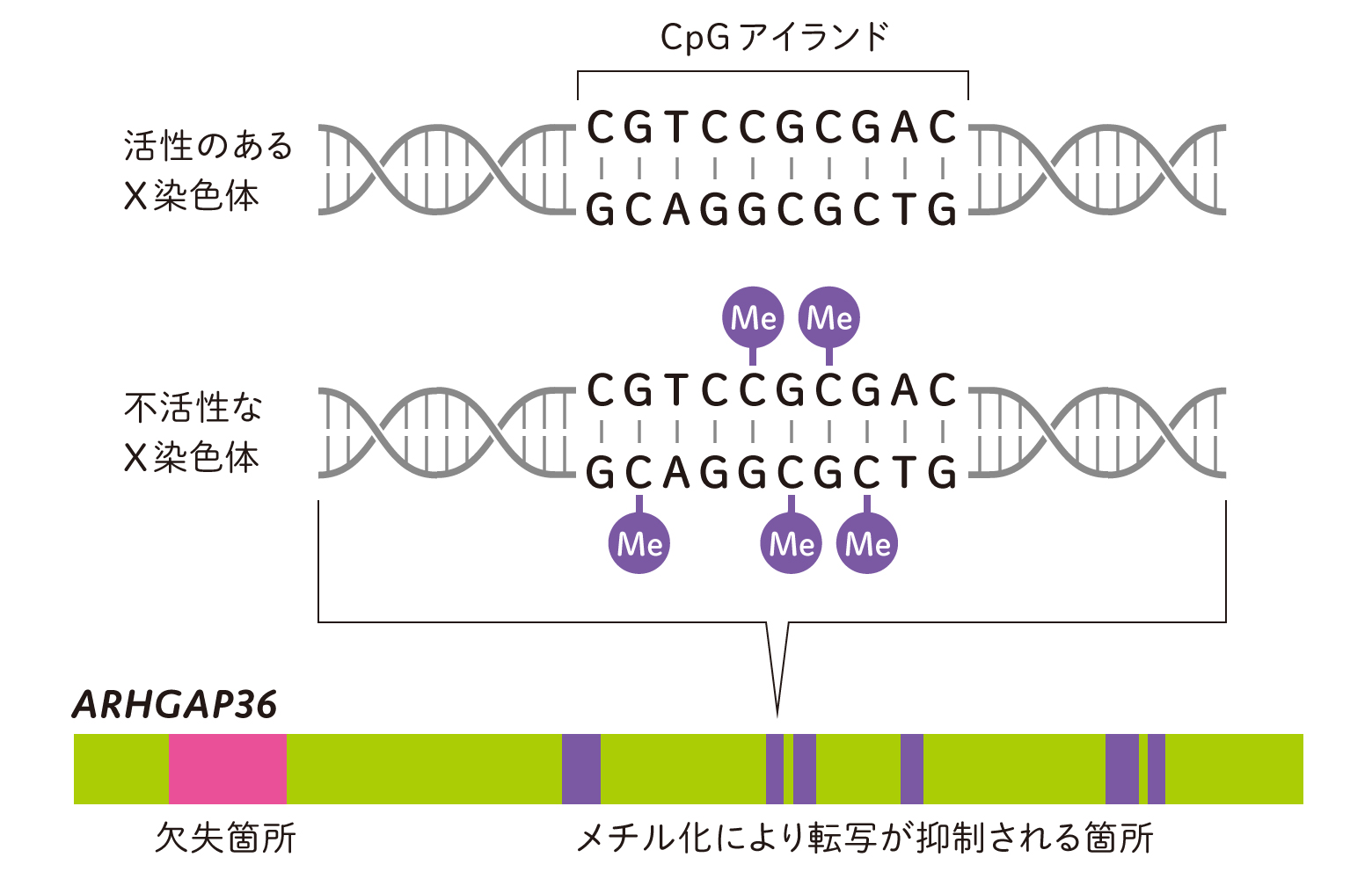
(図4)ネコARHGAP36遺伝子のメスに特異的なメチル化
DNAメチル化状態を次世代シーケンサーにより解析したところ、 メスでのみARHGAP36遺伝子のCpGアイランド(紫色部分)に中等度(2本のX染色体のうち1本)のメチル化(Me)が確認された。
このようにメスの細胞の中に2本あるX染色体のうちどちらか1本は不活性化されます。これによりX染色体を1本しかもたないオスとの間で遺伝子の発現量を均等にしているのだと考えられています。メスの2本のX染色体のどちらが不活性化するかは、個体発生において、神経堤細胞から色素細胞へと分化していく過程で細胞ごとにランダムに決まり、その後は、その不活性化状態を維持したまま領域を占めるように増殖します。そのため、ある程度の広がりをもって黒毛、オレンジ毛の領域が生まれ、三毛猫の場合は色素細胞が到達しなかった白毛の領域も加わって、全体として、3色のまだら模様が描き出されるのだと考えられます。
X染色体不活性化は、X・Y性染色体をもつ哺乳動物に共通のしくみで、ヒトの女性でも起きています。ARHGAP36遺伝子も哺乳動物が共通にもつ遺伝子ですが、毛色の決定に関わるという機能は、今のところイエネコでしか確認されていません。マウス・ヒト・イエネコはゲノム上に同じ遺伝子をもちながら、その働き方を種ごとに柔軟に変える―共通のものから多様を生み出す進化のしくみを、ここにも見ることができるのではないでしょうか。
5. 付かず離れずのネコとヒト
私たちの研究は多くの愛猫家の皆さんに支えられました。実は、この研究のスタート時点で私は定年退職を迎えましたので、クラウドファンディングを通して研究目標を社会へ公開し、資金面でのご支援をいただきました。その際、多くの応援メッセージをもらえたことが、研究を進める上で大きな励みとなりました。サンプル提供や情報交換についても、一般の方々がすすんで協力してくださり、小さいながらも「シチズン・サイエンス」を実践できたことを嬉しく思っています。また研究目標を開示し、成果をいち早く共有したことで、様々な分野の先生たちとの相乗効果を得る「オープン・イノベーション」を実践できたようにも思います。
冒頭で述べたように、誰も取り組まないならば自分たちでと始めた研究でしたが、途中で、同じテーマに取り組んでいる研究グループがあるらしい、との噂が聞こえてきました。スタンフォード大学のグレッグ・バース先生の研究グループでした。彼らの専門分野は色素の生物学で、私たちはエピジェネティクス。専門分野が違うので、お互いに競合するのでなく協力する方がメリットがあると考えて、「情報交換しよう」と連絡したところ、即 “O K ” と返ってきました。早速、オンラインで話し合って、双方の論文を同時投稿することにしました。独立に行っていた二つの研究が同じ結果を示したことで、信憑性も高まり、評価は上々、「オレンジ遺伝子解明」のニュースが世界を駆け巡りました。
イエネコのゲノムデータを確認すると、アメリカ・ヨーロッパ・アフリカ・アジアなど、世界中のオレンジ毛のネコがまったく同じ欠失をもっているので、オレンジ毛をもつネコの起源は一つではないかと考えられます。これについては、科学ジャーナリストの方が私たちの論文に寄せて書いた記事の中で、ネコの遺伝学の専門家であるレスリー・ライオン先生が、「エジプトにはオレンジ色のネコのミイラがあるようだ」と述べてくださっています。骨だと毛色はわかりませんが、ミイラなら毛の色を確認できるかもしれません。いろいろな生きものが描き込まれた古代エジプトの壁画「狩猟の図」(図5)には、オレンジ毛のネコの姿も見えます。もしも、ミイラのゲノムが解読できれば、3千年以上前まで遡って確認することができるかもしれないので、なんとか実現できないかと、可能性を探り始めたところです。
イエネコは、今から1万年から8千年ほど前に、リビアヤマネコから分岐したと考えられています。生まれは西アジアから北アフリカにかけての中近東で、ヒトの歴史では農耕が始まった頃に重なります。イエネコは野生動物ではありませんし、狩猟採集時代からヒトの伴侶動物だったイヌとも生態が異なります。「付かず離れず」というヒトとネコとの関係が、もうこの頃から始まっていたかもしれません。こうしていろいろと思案していると、ネコとのご縁は、これからもまだまだ続きそう、いや深まっていきそうな気がします。

(図5)「狩猟の図」所蔵:大英博物館

佐々木 裕之
(ささき・ひろゆき)
1956年福岡市生まれ。1987年九州大学大学院医学系研究科修了(医学博士)。1990年英国ケンブリッジのAFRC研究所及びウェルカム/CRC研究所で、ゲノムインプリンティングを発見したアジム・スラーニ博士とエピジェネティクス研究に携わる。国立遺伝学研究所教授、総合研究大学院大学教授、九州大学生体防御医学研究所教授・所長、九州大学副学長、九州大学高等研究院研究院長を歴任。定年退職後の現在もクラウドファンドなどを活用し、精力的に市民や社会と共創するサイエンスを実践している。著書に『エピジェネティクス』(編著,シュプリンガー・フェアラーク東京)『エピジェネティクス入門ー三毛猫の模様はどう決まるのかー』(岩波書店)など。日本人類遺伝学会学会賞(2009)、日本遺伝学会・木原賞(2012)、紫綬褒章(2015)、上原賞(2018)、科学技術分野の文部科学大臣表彰 科学技術賞受賞(2026)。
.jpg)

.jpg)
.jpg)










