

顧問の西川伸一を中心に館員が、今進化研究がどのようにおこなわれているかを紹介していきます。進化研究とは何をすることなのか? 歴史的背景も含めお話しします。
バックナンバー
失う進化
2015年3月16日
進化というと新しい形質を獲得することに頭が行きがちだが、必要ないものを失う進化もある。さらに、必要ないのに捨てることができず、進化の自由を制限する形質もある。2回にわたって、進化は獲得するばかりでないことを示す例について見ていこう。とはいえ、この話題に近い話はすでに第25話に出てきている。すなわち、陸上哺乳類が水中生活を始めることで、邪魔な長い四肢を、泳ぎに適したヒレ足に変化させた例だ。図1の鯨の骨格を見てほしい。
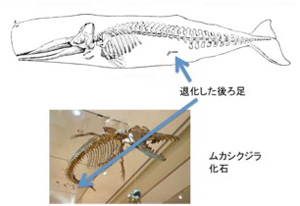
図1 鯨の骨格図とムカシクジラの化石。矢印は退化した後ろ足。(Wikimedia Commonsより)
前のヒレ足の骨格を見ると、ゲーテが原型の概念を用いて説明したよう(第26話参照)に、陸棲哺乳類の持つ前足の骨と相同の骨が形を変化させ、時には融合することで、足からヒレ足への形態変化が起こっている。ただ外から見ると鯨には後ろのヒレ足はない。しかし骨格で見ると、矢印で示した小さな骨のかけらが残っており、実際には退化したことがわかる。このことは、今から5千万年前に生息した鯨の先祖ムカシクジラ化石にしっかりとした足が残っていることからもわかる。このようにクジラ類は5千万年の進化の過程で、必要なくなった後ろ足の骨格をほぼ捨て去ることに成功している。それでも、申し訳のように骨のかけらが出来てしまうのは不思議だ。これは発生過程で四肢を誘導する仕組みを残したまま、足の骨の形成過程を抑制することで、後ろ足を退化させているためだ。実際イルカ胎児を見ると(図2)、肢芽と呼ばれる構造が誘導されていることがわかる。発生過程をおった研究から、一旦誘導された肢芽がそれ以上発達できない過程を見ることができる(Thewissen et al, Developmental basis for hind limb loss in dolphin and origin of the cetacean body plan, Proc Natl Acad Sci USA, 103, 8414, 2006に詳しい:ダウンロード自由)。このように、一旦苦労して獲得した仕組みは、そう簡単に捨てられない。
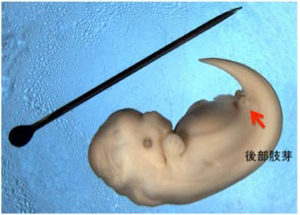
図2:5週齢のイルカ胎児。ピンの長さは1インチ。(Wikimedia Commonsより)
四肢と同様に、脊椎動物の上陸作戦を考えるとき最初に頭に浮かぶのが、エラから肺呼吸への進化だが、では水中生活に応じてこの逆は起こるのだろうか?残念ながら、一旦肺呼吸が進化した動物で、エラ呼吸が復活する例はなさそうだ。すべての水棲哺乳動物は、どんなに長時間潜水ができても、肺を使っているのでいつかは表面に浮かんできて空気を取り入れる。クジラの潮吹きはまさにこの時に起こる。長時間の潜水は、呼吸システムの変化ではなく、心筋や酸素消費システムを変化させ、長時間の潜水を可能にしている(第25話)。どうして戻れないのか?金輪際戻れないのか?確かな答はないが、戻るのが簡単でないことは想像できる。呼吸は生命にとって必須の機能で、水中と陸上の両方で生きるためには、肺魚のように、2重の呼吸システムを持つ中間段階が開発できて初めて、新しいシステムへの移行が可能になる。もし陸上哺乳動物がエラ呼吸に逆戻りする可能性があるなら、肺魚と同じように、肺呼吸とエラ呼吸の両方が並立する段階が必要だろう。ただ、たとえこれが可能だったとしても、空気中と比べると水中の酸素濃度は低く、肺の代わりにエラを使うには酸素摂取効率が低すぎるだろう。おそらく、肺呼吸を使い続けたほうが、より高い身体能力を維持できるはずだ。このようにほとんどの進化の道筋は高い機能を達成するため袋小路に入ってしまって、環境が変わってもそう簡単に逆戻りはできない。
では陸棲から水棲への生活圏の移行に伴い、脊椎動物の上陸作戦で見られたのと逆のことが起こったような例はあるのだろうか?一見この条件に合致した逆向き進化の例が、臭い物質と結合する嗅覚受容体遺伝子に見られることが知られている。嗅覚受容体は、空気中に存在する微量なニオイ物質(多くは脂溶性)と結合することで、嗅覚細胞の神経興奮を誘導する受容体で、鼻粘膜に発現している(図3)。この分子がニオイ物質と結合するとGタンパクを介してアデニルシクラーゼを活性化し、cAMPを介してナトリウムチャンネルを開いて、神経の興奮を誘導する。
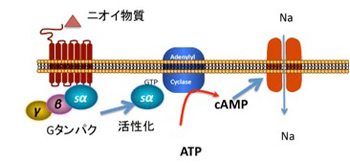
図3 嗅覚受容体シグナル。ニオイ物質に結合するとGタンパクが活性化され、GDP-GTP変換が起こり、これがアデニルシクラーゼを活性化する。作られたcAMPが次にナトリウムチャンネルを開き、細胞を興奮させる。
この分子をコードする遺伝子は、1991年リンダ・バック(Linda B. Buck)とリチャード・アクセル(Richard Axel)によりクローニングされ、その後嗅覚についての研究は大きく進展した。バックとアクセルはニオイ受容体および嗅覚系組織の発見で2004年、ノーベル医学生理学賞を受賞する。このおかげで、私たちは何百もの嗅覚受容体遺伝子をゲノムに持っていることがわかっている。哺乳動物では300−2000種類の嗅覚受容体が存在することが知られているが、犯人のニオイを嗅ぎ当てることができるほど高感度な警察犬の能力を考えると、これでも少ないのではと心配する。さて、ゲノム上の嗅覚受容体遺伝子の数を脊椎動物で比べてみると、硬骨魚(多い方のゼブラフィッシュで約150個)から両生類への進化過程で(アフリカツメガエル約1000個)、数が上昇することがわかっている。この傾向から、水中ではそれほど微妙なニオイを認識する必要がないが、上陸後は食物や天敵の発見まで、行動決定に対するニオイの影響が大きくなっていくことが想像される。では、陸棲から水棲へ移行した哺乳動物では嗅覚受容体の減少が見られるのだろうか?実際、鯨の生態を考えてみると、揮発性のにおいで餌や天敵を見つける必要はなくなっているように思える。必要の無くなった嗅覚受容体遺伝子は消失したのだろうか?つい先ごろ京大理学部から日本動物学会発行の雑誌Zoological Lettersに掲載された論文(Zoological letters 1、9,2015)を元にこの問題を調べてみよう。この研究では、京大グループが解読したクロミンククジラのゲノムデータと、データベースから得られる牛及びバンドウイルカの嗅覚受容体を比べている。

図4 機能的嗅覚受容体遺伝子数の比較。Zoological letters 1、9,2015を改変して転載。
図4はこの論文の図2に掲載されているデータを改変して転載した。牛は陸棲哺乳動物の中では、もっともイルカや鯨と近縁で、5−6千万年前に牛と別れたと考えられるが(第25話参照)、この間に機能的嗅覚受容体の数が大きく減少していることがわかる。この論文では嗅神経が投射する嗅球の構造についても調べ、水棲への移行が始まってすぐに、嗅球上部構造が消失し始め、ハクジラになると嗅球自体がなくなっていることを示している。陸上に暮らしていても、1000を越す嗅覚受容体遺伝子を持つマウスと、たかだか300しか持たない人間の嗅球を比べると、図5にあるように脳内での存在感が全く違う。
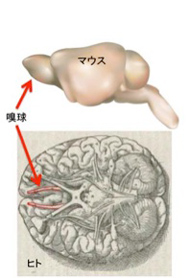
図5 マウスとヒトの嗅球の比較。脳全体から見た比率で言うと、ヒト嗅球は退化していると言える。(Wikimedia Commonsより)
これを見ると、他の器官と比べた時、神経や脳は必要に合わせて極めて獲得や消失が簡単なことがわかる。視覚についても同じことが言える。動物種によっては色が見えなかったりするのはよく知られた事実で、色認識に関わるフォトプシンの進化は獲得や消失が進化の過程で何回も起こっている。このような出没自在なシステムでは、上陸作戦と水棲への移行で逆のプロセスが起こるのを確かに観察できる。次回は、低温に適応して遺伝子が失われる例を紹介する。